

A CRISPR Screen Reveals Resistance Mechanisms to CD3-Bispecific Antibody Therapy
- PMID: 33177106
- PMCID: PMC8601150
- DOI: 10.1158/2326-6066.CIR-20-0080
A CRISPR Screen Reveals Resistance Mechanisms to CD3-Bispecific Antibody Therapy
Abstract
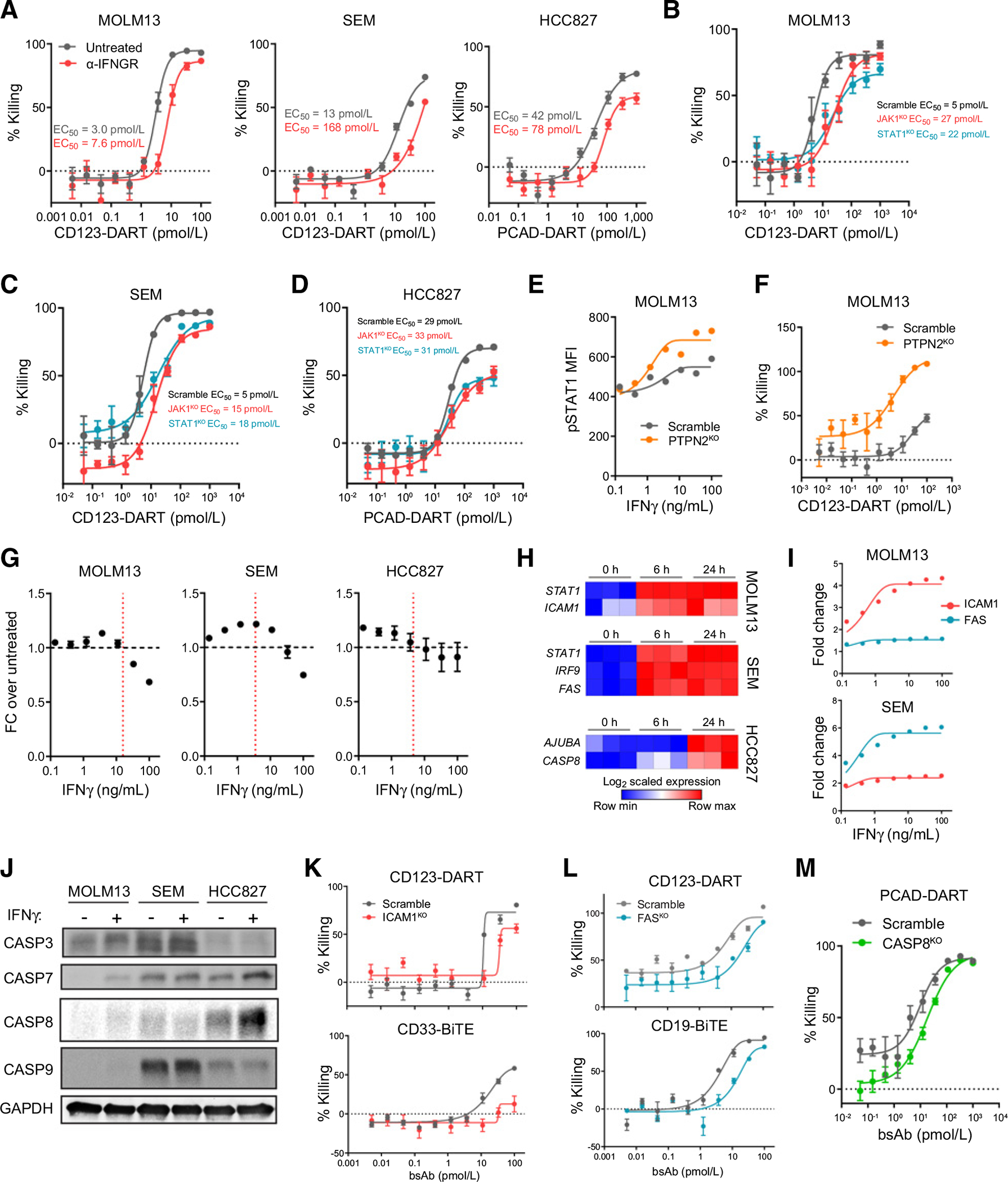
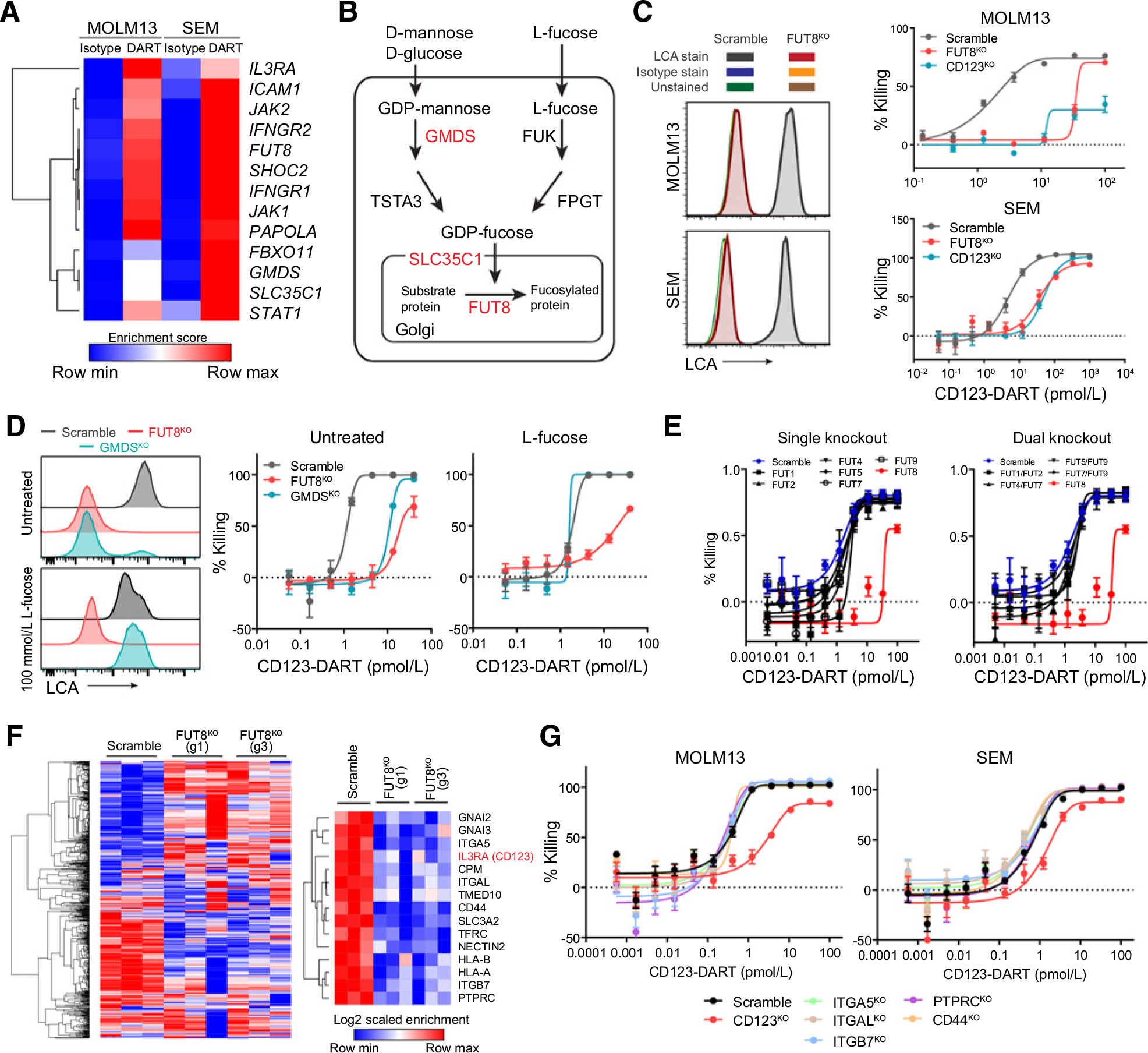
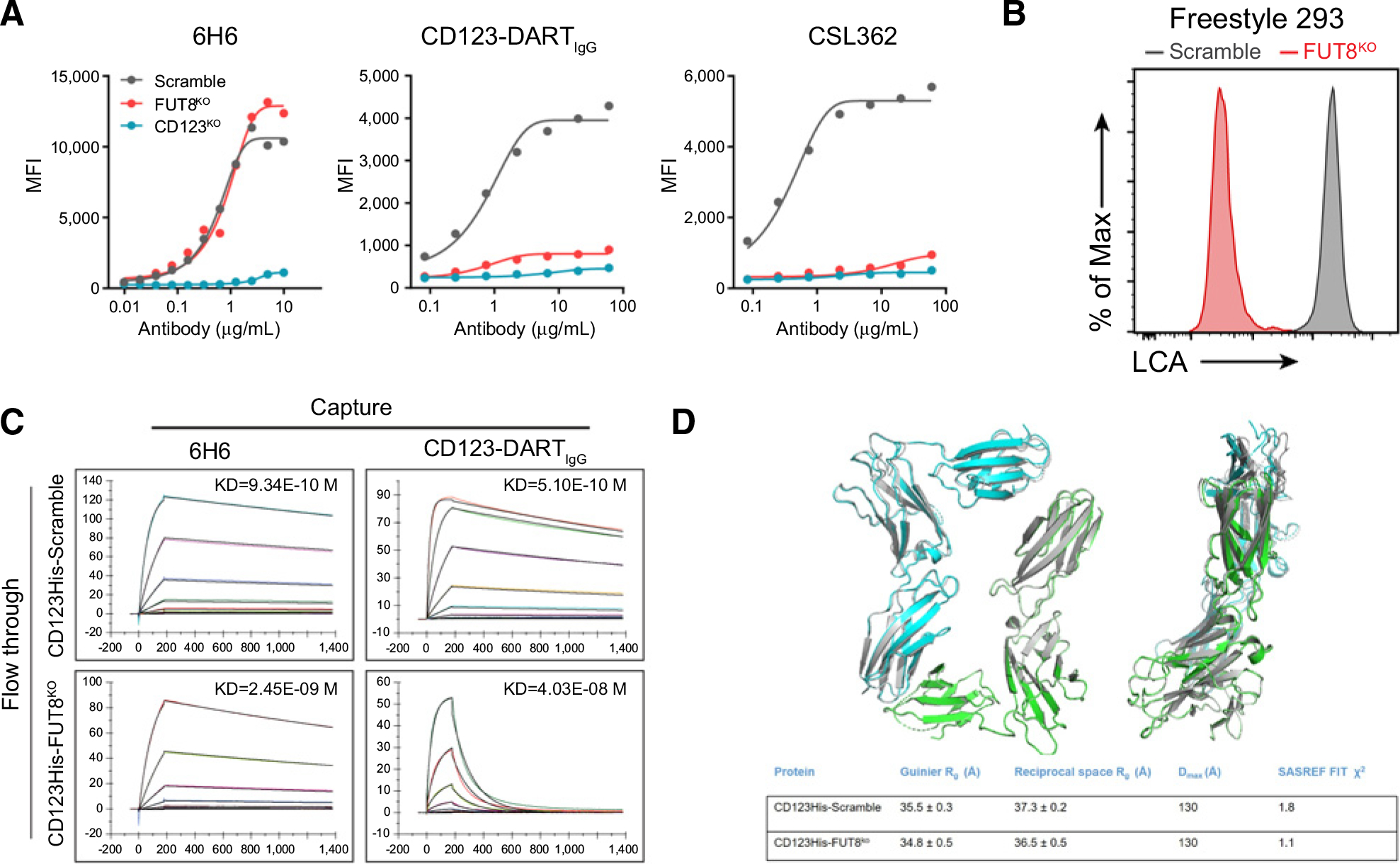
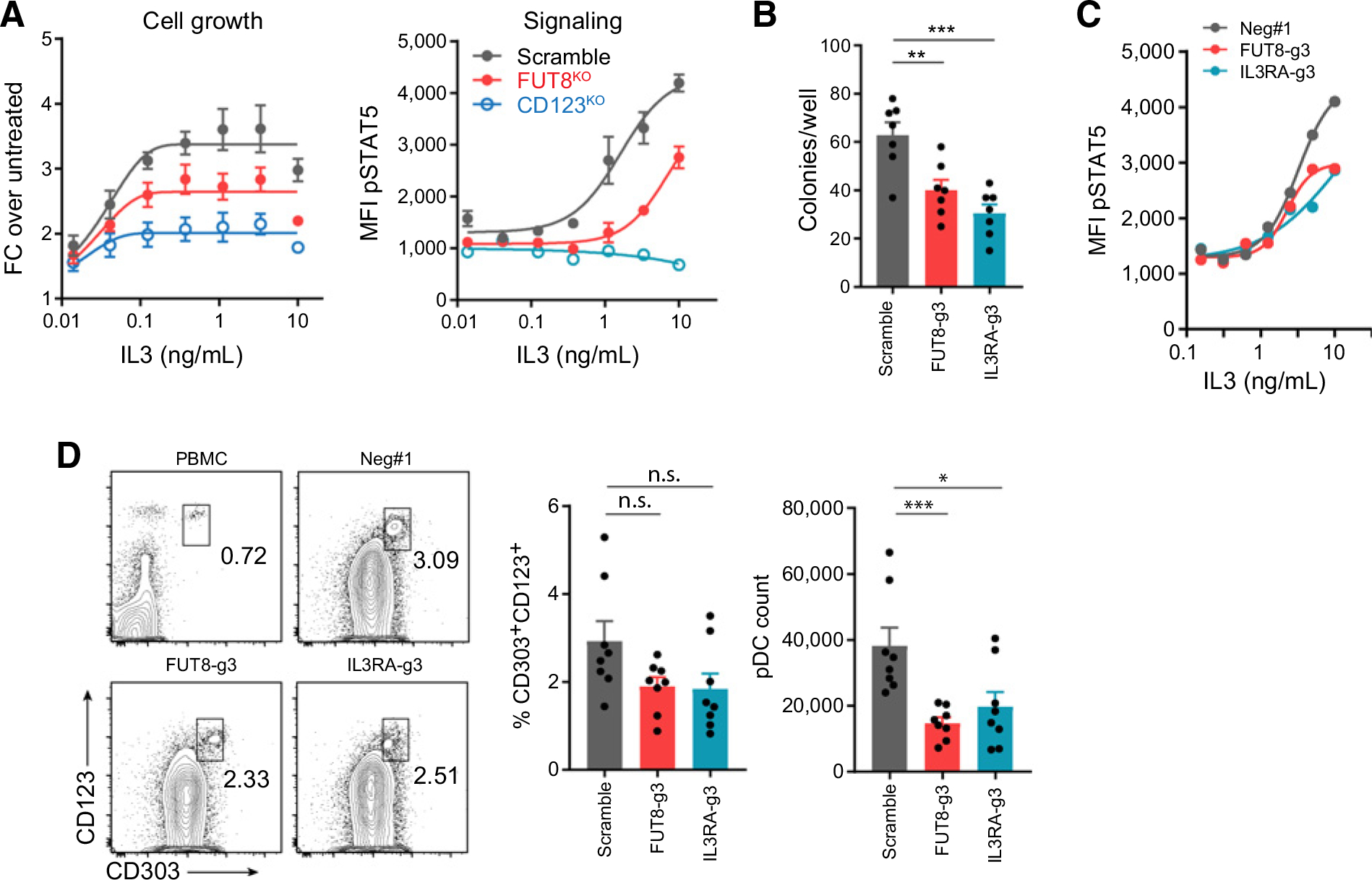
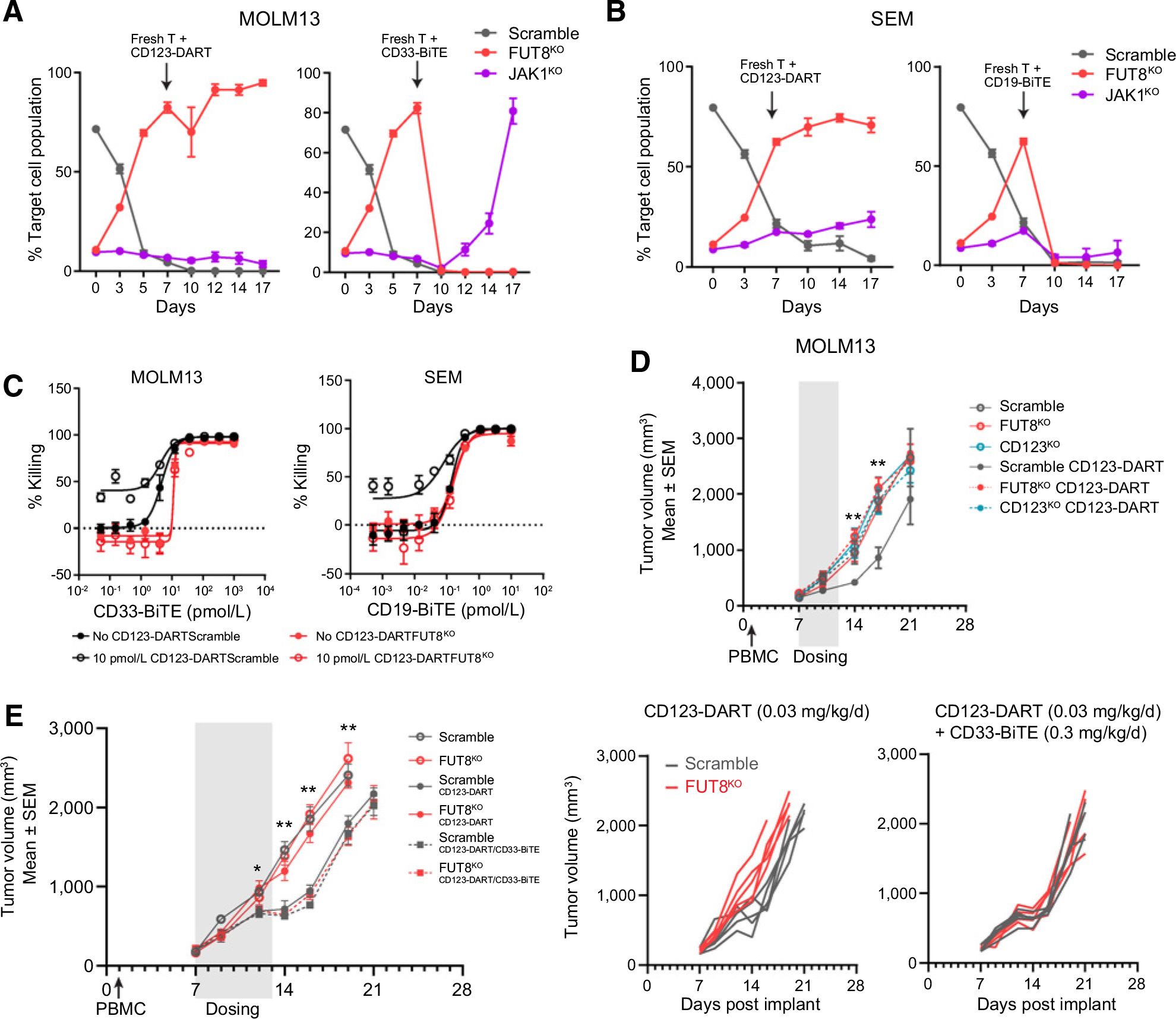
CD3-bispecific antibodies represent an important therapeutic strategy in oncology. These molecules work by redirecting cytotoxic T cells to antigen-bearing tumor cells. Although CD3-bispecific antibodies have been developed for several clinical indications, cases of cancer-derived resistance are an emerging limitation to the more generalized application of these molecules. Here, we devised whole-genome CRISPR screens to identify cancer resistance mechanisms to CD3-bispecific antibodies across multiple targets and cancer types. By validating the screen hits, we found that deficiency in IFNγ signaling has a prominent role in cancer resistance. IFNγ functioned by stimulating the expression of T-cell killing-related molecules in a cell type-specific manner. By assessing resistance to the clinical CD3-bispecific antibody flotetuzumab, we identified core fucosylation as a critical pathway to regulate flotetuzumab binding to the CD123 antigen. Disruption of this pathway resulted in significant resistance to flotetuzumab treatment. Proper fucosylation of CD123 was required for its normal biological functions. In order to treat the resistance associated with fucosylation loss, flotetuzumab in combination with an alternative targeting CD3-bispecific antibody demonstrated superior efficacy. Together, our study reveals multiple mechanisms that can be targeted to enhance the clinical potential of current and future T-cell-engaging CD3-bispecific antibody therapies.
©2020 American Association for Cancer Research.
Figures

References
-
- Clynes RA, Desjarlais JR. Redirected T cell cytotoxicity in cancer therapy. Annu Rev Med 2019;70:437–50. - PubMed
-
- Uy GL, Godwin J, Rettig MP, Vey N, Foster M, Arellano ML, et al. Preliminary results of a phase 1 study of flotetuzumab, a CD123 × CD3 bispecific dart (R) protein, in patients with relapsed/refractory acute myeloid leukemia and myelodysplastic syndrome. Blood 2017;130 Suppl 1:637.
-
- Aldoss I, Song J, Stiller T, Nguyen T, Palmer J, O’Donnell M, et al. Correlates of resistance and relapse during blinatumomab therapy for relapsed/refractory acute lymphoblastic leukemia. Am J Hematol 2017;92: 858–65. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
