

SARS-CoV-2 receptor is co-expressed with elements of the kinin-kallikrein, renin-angiotensin and coagulation systems in alveolar cells
- PMID: 33177594
- PMCID: PMC7658217
- DOI: 10.1038/s41598-020-76488-2
SARS-CoV-2 receptor is co-expressed with elements of the kinin-kallikrein, renin-angiotensin and coagulation systems in alveolar cells
Abstract
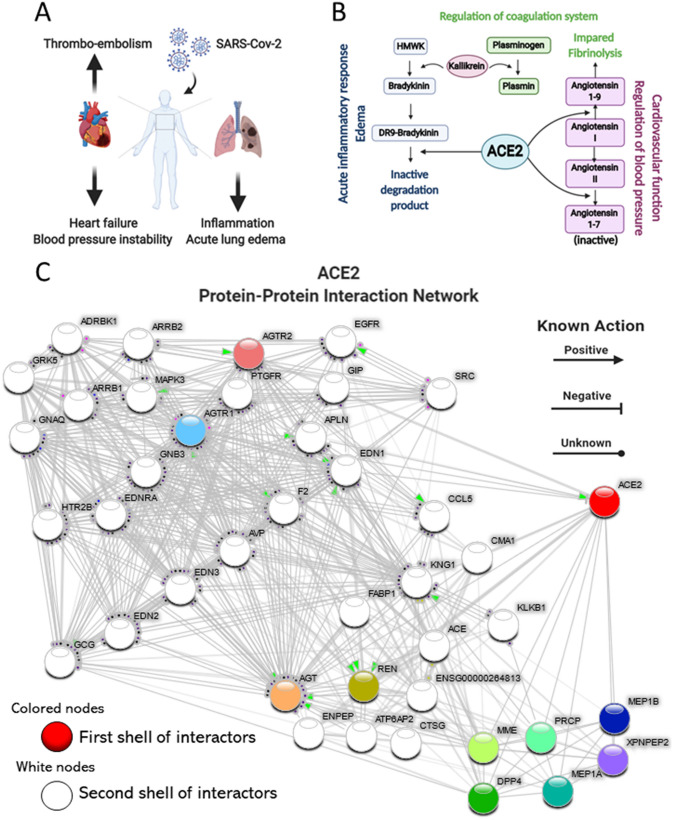
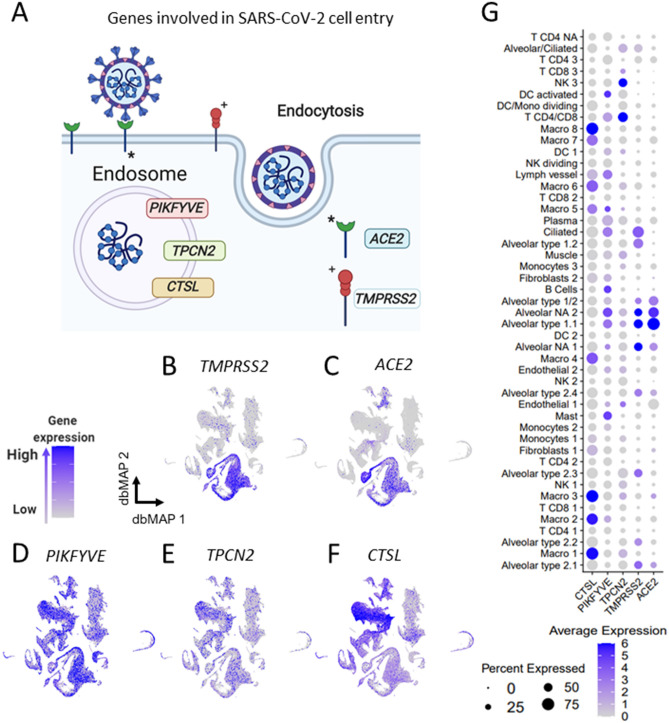
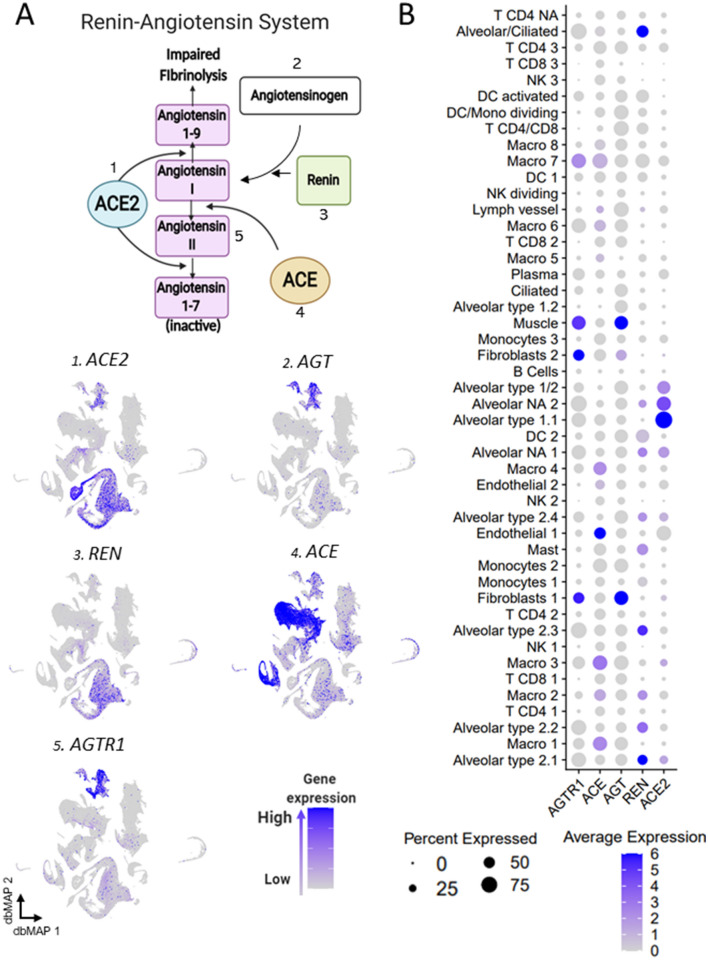
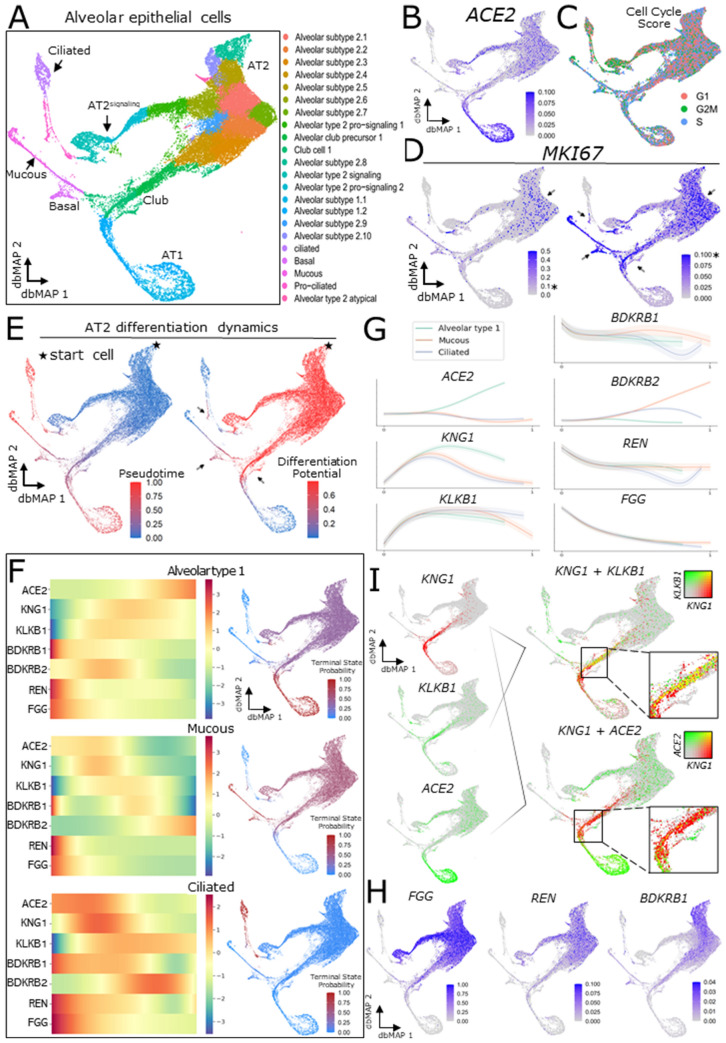
SARS-CoV-2, the pathogenic agent of COVID-19, employs angiotensin converting enzyme-2 (ACE2) as its cell entry receptor. Clinical data reveal that in severe COVID-19, SARS-CoV-2 infects the lung, leading to a frequently lethal triad of respiratory insufficiency, acute cardiovascular failure, and coagulopathy. Physiologically, ACE2 plays a role in the regulation of three systems that could potentially be involved in the pathogenesis of severe COVID-19: the kinin-kallikrein system, resulting in acute lung inflammatory edema; the renin-angiotensin system, promoting cardiovascular instability; and the coagulation system, leading to thromboembolism. Here we assembled a healthy human lung cell atlas meta-analysis with ~ 130,000 public single-cell transcriptomes and show that key elements of the bradykinin, angiotensin and coagulation systems are co-expressed with ACE2 in alveolar cells and associated with their differentiation dynamics, which could explain how changes in ACE2 promoted by SARS-CoV-2 cell entry result in the development of the three most severe clinical components of COVID-19.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
