

Persistent transcriptional programmes are associated with remote memory
- PMID: 33177708
- PMCID: PMC9097329
- DOI: 10.1038/s41586-020-2905-5
Persistent transcriptional programmes are associated with remote memory
Erratum in
-
Author Correction: Persistent transcriptional programmes are associated with remote memory.Nature. 2025 Aug;644(8077):E37. doi: 10.1038/s41586-025-09463-4. Nature. 2025. PMID: 40770108 No abstract available.
Abstract
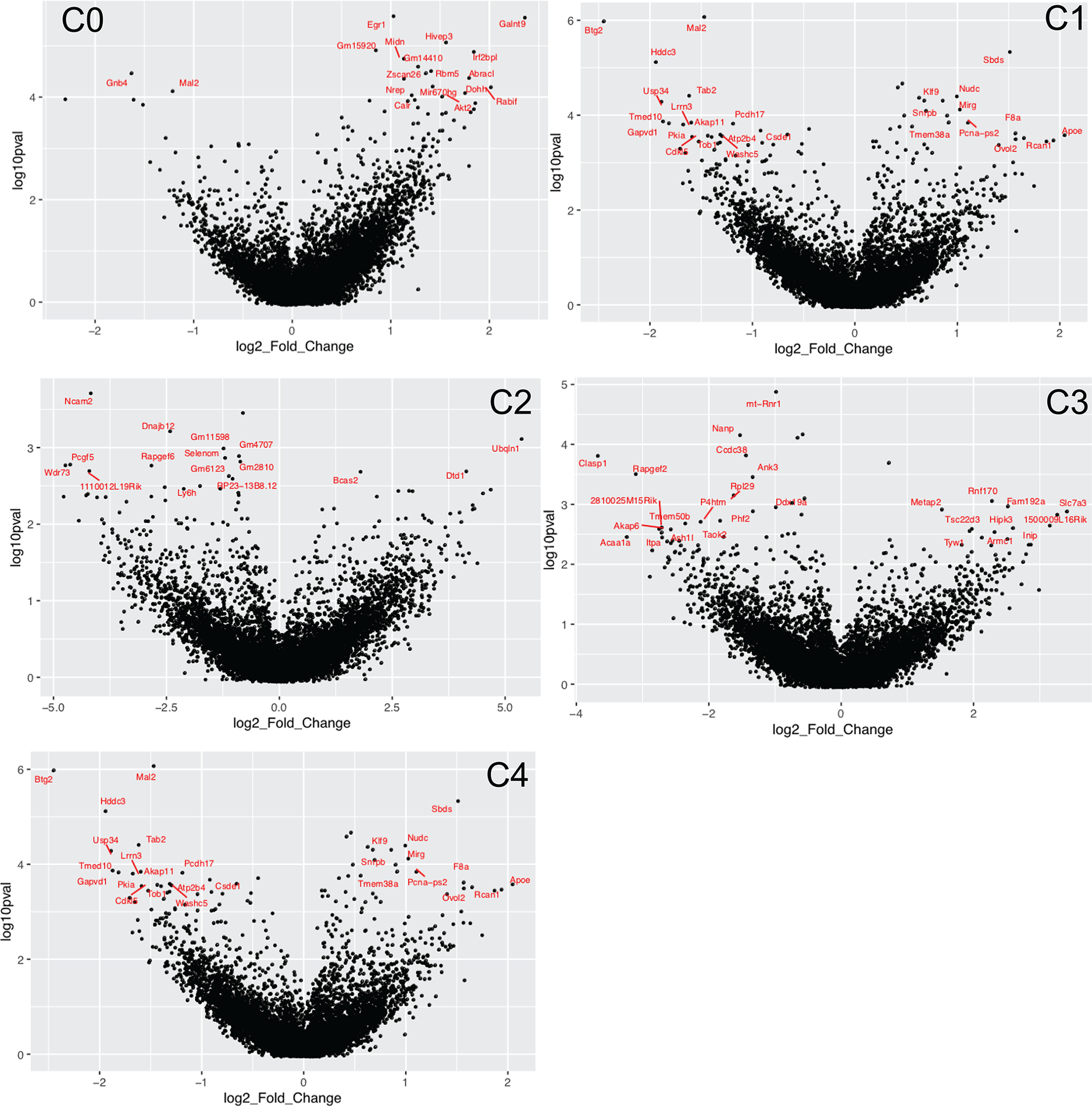
The role of gene expression during learning and in short-term memories has been studied extensively1-3, but less is known about remote memories, which can persist for a lifetime4. Here we used long-term contextual fear memory as a paradigm to probe the single-cell gene expression landscape that underlies remote memory storage in the medial prefrontal cortex. We found persistent activity-specific transcriptional alterations in diverse populations of neurons that lasted for weeks after fear learning. Out of a vast plasticity-coding space, we identified genes associated with membrane fusion that could have important roles in the maintenance of remote memory. Unexpectedly, astrocytes and microglia also acquired persistent gene expression signatures that were associated with remote memory, suggesting that they actively contribute to memory circuits. The discovery of gene expression programmes associated with remote memory engrams adds an important dimension of activity-dependent cellular states to existing brain taxonomy atlases and sheds light on the elusive mechanisms of remote memory storage.
Conflict of interest statement
Figures














References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
