

Dissecting the Genetic and Etiological Causes of Primary Microcephaly
- PMID: 33178111
- PMCID: PMC7593518
- DOI: 10.3389/fneur.2020.570830
Dissecting the Genetic and Etiological Causes of Primary Microcephaly
Abstract
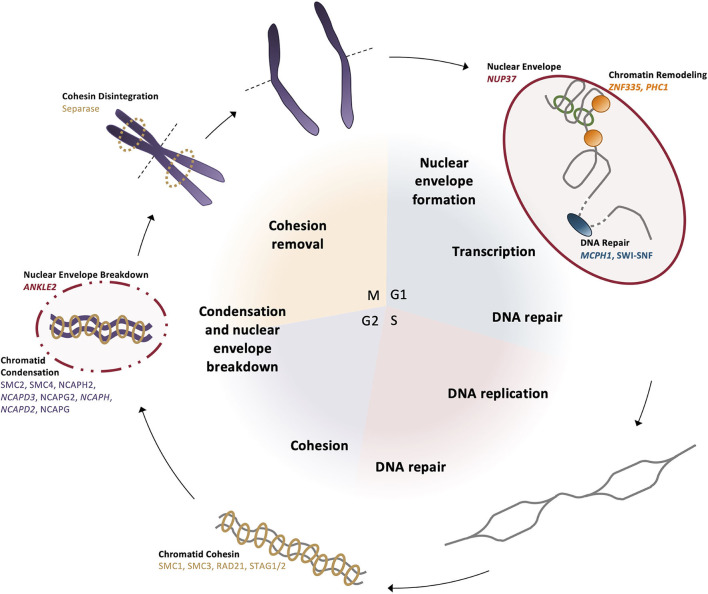
Autosomal recessive primary microcephaly (MCPH; "small head syndrome") is a rare, heterogeneous disease arising from the decreased production of neurons during brain development. As of August 2020, the Online Mendelian Inheritance in Man (OMIM) database lists 25 genes (involved in molecular processes such as centriole biogenesis, microtubule dynamics, spindle positioning, DNA repair, transcriptional regulation, Wnt signaling, and cell cycle checkpoints) that are implicated in causing MCPH. Many of these 25 genes were only discovered in the last 10 years following advances in exome and genome sequencing that have improved our ability to identify disease-causing variants. Despite these advances, many patients still lack a genetic diagnosis. This demonstrates a need to understand in greater detail the molecular mechanisms and genetics underlying MCPH. Here, we briefly review the molecular functions of each MCPH gene and how their loss disrupts the neurogenesis program, ultimately demonstrating that microcephaly arises from cell cycle dysregulation. We also explore the current issues in the genetic basis and clinical presentation of MCPH as additional avenues of improving gene/variant prioritization. Ultimately, we illustrate that the detailed exploration of the etiology and inheritance of MCPH improves the predictive power in identifying previously unknown MCPH candidates and diagnosing microcephalic patients.
Keywords: cell cycle; genetics; microcephaly; neurogenesis; rare disease (RD).
Copyright © 2020 Jean, Stuart and Tarailo-Graovac.
Figures
Similar articles
-
Comprehensive review on the molecular genetics of autosomal recessive primary microcephaly (MCPH).Genet Res (Camb). 2018 Aug 8;100:e7. doi: 10.1017/S0016672318000046. Genet Res (Camb). 2018. PMID: 30086807 Free PMC article. Review.
-
Identification of Pathogenic Mutations in Primary Microcephaly- (MCPH-) Related Three Genes CENPJ, CASK, and MCPH1 in Consanguineous Pakistani Families.Biomed Res Int. 2022 Mar 3;2022:3769948. doi: 10.1155/2022/3769948. eCollection 2022. Biomed Res Int. 2022. PMID: 35281599 Free PMC article.
-
What next-generation sequencing (NGS) technology has enabled us to learn about primary autosomal recessive microcephaly (MCPH).Mol Cell Probes. 2015 Oct;29(5):271-81. doi: 10.1016/j.mcp.2015.05.015. Epub 2015 Jun 4. Mol Cell Probes. 2015. PMID: 26050940 Review.
-
Molecular Genetics of Microcephaly Primary Hereditary: An Overview.Brain Sci. 2021 Apr 30;11(5):581. doi: 10.3390/brainsci11050581. Brain Sci. 2021. PMID: 33946187 Free PMC article. Review.
-
Autosomal Recessive Primary Microcephaly (MCPH): clinical manifestations, genetic heterogeneity and mutation continuum.Orphanet J Rare Dis. 2011 Jun 13;6:39. doi: 10.1186/1750-1172-6-39. Orphanet J Rare Dis. 2011. PMID: 21668957 Free PMC article. Review.
Cited by
-
The role of histone methyltransferases in neurocognitive disorders associated with brain size abnormalities.Front Neurosci. 2023 Feb 10;17:989109. doi: 10.3389/fnins.2023.989109. eCollection 2023. Front Neurosci. 2023. PMID: 36845425 Free PMC article. Review.
-
Congenital Microcephaly: A Debate on Diagnostic Challenges and Etiological Paradigm of the Shift from Isolated/Non-Syndromic to Syndromic Microcephaly.Cells. 2023 Feb 16;12(4):642. doi: 10.3390/cells12040642. Cells. 2023. PMID: 36831309 Free PMC article. Review.
-
AUTS2 Syndrome: Molecular Mechanisms and Model Systems.Front Mol Neurosci. 2022 Mar 31;15:858582. doi: 10.3389/fnmol.2022.858582. eCollection 2022. Front Mol Neurosci. 2022. PMID: 35431798 Free PMC article. Review.
-
Autophagy in neural stem cells and glia for brain health and diseases.Neural Regen Res. 2024 Apr;19(4):729-736. doi: 10.4103/1673-5374.382227. Neural Regen Res. 2024. PMID: 37843206 Free PMC article. Review.
-
The Multiple Mitotic Roles of the ASPM Orthologous Proteins: Insight into the Etiology of ASPM-Dependent Microcephaly.Cells. 2023 Mar 16;12(6):922. doi: 10.3390/cells12060922. Cells. 2023. PMID: 36980263 Free PMC article. Review.
References
Publication types
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous