

Human de novo purine biosynthesis
- PMID: 33179964
- PMCID: PMC7869020
- DOI: 10.1080/10409238.2020.1832438
Human de novo purine biosynthesis
Abstract
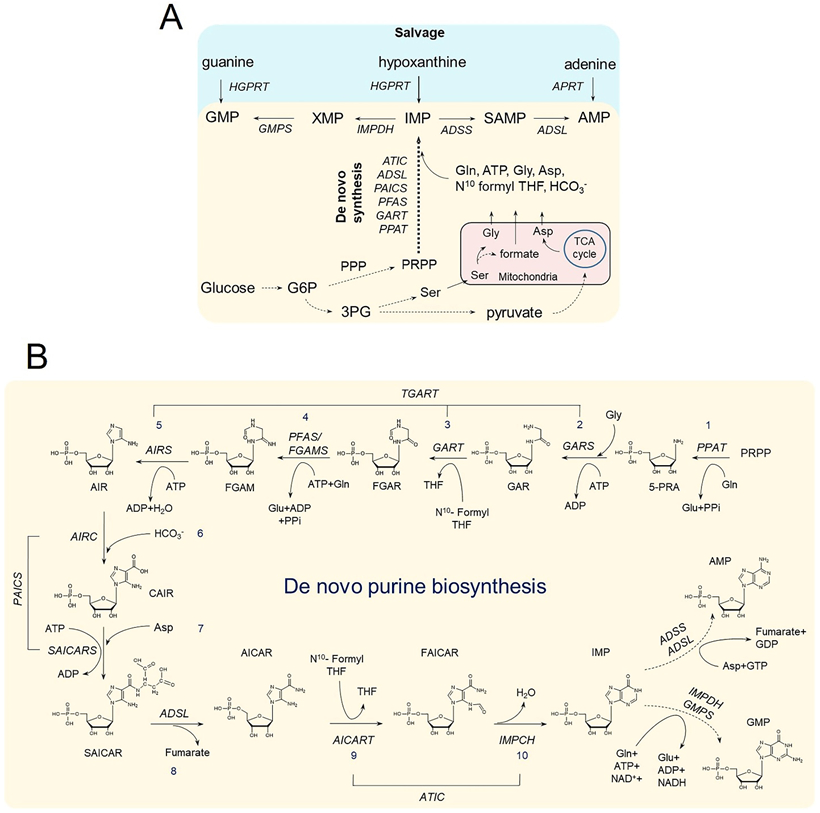
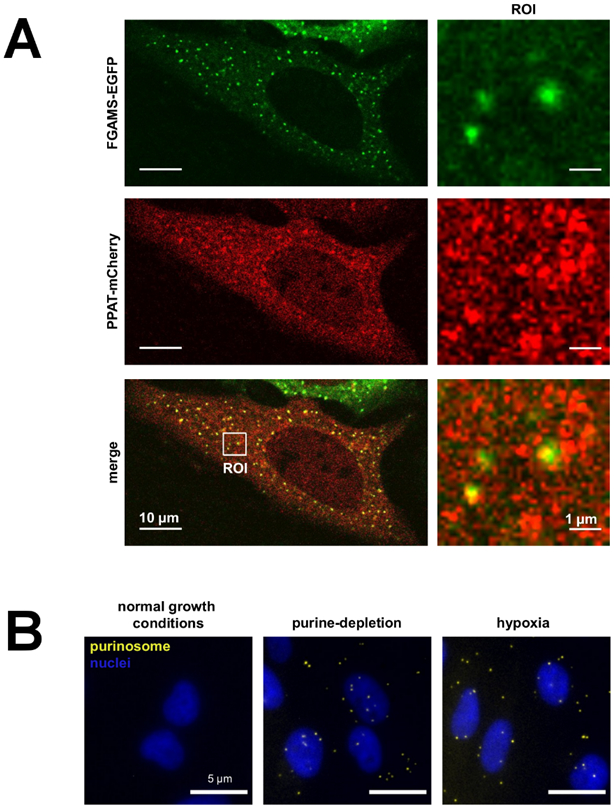
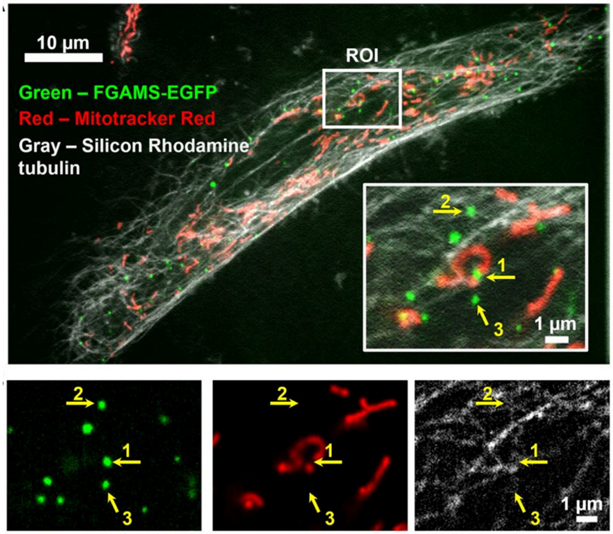
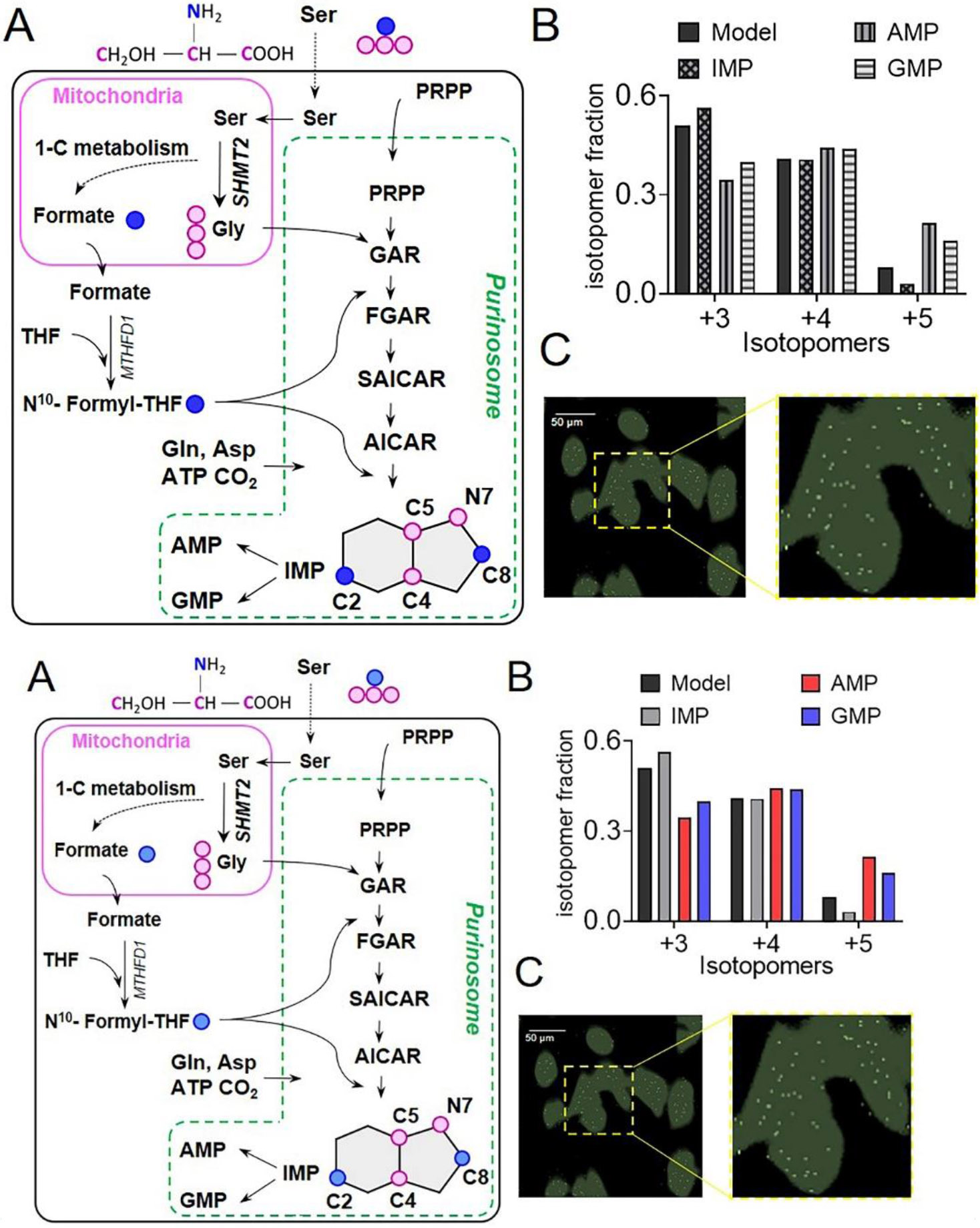
The focus of this review is the human de novo purine biosynthetic pathway. The pathway enzymes are enumerated, as well as the reactions they catalyze and their physical properties. Early literature evidence suggested that they might assemble into a multi-enzyme complex called a metabolon. The finding that fluorescently-tagged chimeras of the pathway enzymes form discrete puncta, now called purinosomes, is further elaborated in this review to include: a discussion of their assembly; the role of ancillary proteins; their locus at the microtubule/mitochondria interface; the elucidation that at endogenous levels, purinosomes function to channel intermediates from phosphoribosyl pyrophosphate to AMP and GMP; and the evidence for the purinosomes to exist as a protein condensate. The review concludes with a consideration of probable signaling pathways that might promote the assembly and disassembly of the purinosome, in particular the identification of candidate kinases given the extensive phosphorylation of the enzymes. These collective findings substantiate our current view of the de novo purine biosynthetic metabolon whose properties will be representative of how other metabolic pathways might be organized for their function.
Keywords: metabolism de novo purine biosynthesis purinosome metabolon substrate channeling condensate signaling.
Conflict of interest statement
DISCLOSURE STATEMENT
No potential conflict of interest was reported by the authors.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources