

A requirement for PAK1 to support mitochondrial function and maintain cellular redox balance via electron transport chain proteins to prevent β-cell apoptosis
- PMID: 33181191
- PMCID: PMC8123936
- DOI: 10.1016/j.metabol.2020.154431
A requirement for PAK1 to support mitochondrial function and maintain cellular redox balance via electron transport chain proteins to prevent β-cell apoptosis
Abstract
Objective: p21 (Cdc42/Rac1) activated Kinase 1 (PAK1) is a candidate susceptibility factor for type 2 diabetes (T2D). PAK1 is depleted in the islets from T2D donors, compared to control individuals. In addition, whole-body PAK1 knock out (PAK1-KO) in mice worsens the T2D-like effects of high-fat diet. The current study tested the effects of modulating PAK1 levels only in β-cells.
Materials/methods: β-cell-specific inducible PAK1 KO (βPAK1-iKO) mice were generated and used with human β-cells and T2D islets to evaluate β-cell function.
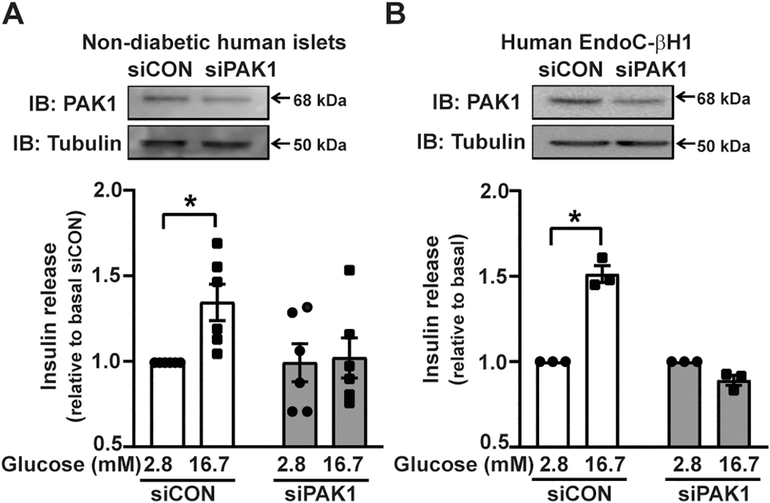
Results: βPAK1-iKO mice exhibited glucose intolerance and elevated β-cell apoptosis, but without peripheral insulin resistance. β-cells from βPAK-iKO mice also contained fewer mitochondria per cell. At the cellular level, human PAK1-deficient β-cells showed blunted glucose-stimulated insulin secretion and reduced mitochondrial function. Mitochondria from human PAK1-deficient β-cells were deficient in the electron transport chain (ETC) subunits CI, CIII, and CIV; NDUFA12, a CI complex protein, was identified as a novel PAK1 binding partner, and was significantly reduced with PAK1 knockdown. PAK1 knockdown disrupted the NAD+/NADH and NADP+/NADPH ratios, and elevated ROS. An imbalance of the redox state due to mitochondrial dysfunction leads to ER stress in β-cells. PAK1 replenishment in the β-cells of T2D human islets ameliorated levels of ER stress markers.
Conclusions: These findings support a protective function for PAK1 in β-cells. The results support a new model whereby the PAK1 in the β-cell plays a required role upstream of mitochondrial function, via maintaining ETC protein levels and averting stress-induced β-cell apoptosis to retain healthy functional β-cell mass.
Keywords: Diabetes; Electron transport chain; Mitochondrial number; Redox imbalance; β-Cell mass.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare no conflicts of interest.
Figures






References
-
- CDC. Centers for disease control and prevention: national diabetes statistics report, 2017. Atlanta, GA: Centers for Disease Control and Prevention, US Department of Health and Human Services; 2017.
-
- Tripathy D, Eriksson KF, Orho-Melander M, Fredriksson J, Ahlqvist G, Groop L. Parallel manifestation of insulin resistance and beta cell decompensation is compatible with a common defect in type 2 diabetes. Diabetologia. 2004;47:782–93. - PubMed
-
- Cohrs CM, Panzer JK, Drotar DM, Enos SJ, Kipke N, Chen C, et al. Dysfunction of persisting β cells is a key feature of early type 2 diabetes pathogenesis. Cell Rep. 2020;31:107469. - PubMed
-
- Butler AE, Janson J, Bonner-Weir S, Ritzel R, Rizza RA, Butler PC. Beta-cell deficit and increased beta-cell apoptosis in humans with type 2 diabetes. Diabetes. 2003;52: 102–10. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- FS/19/39/34447/BHF_/British Heart Foundation/United Kingdom
- PG/14/71/31063/BHF_/British Heart Foundation/United Kingdom
- R01 DK102233/DK/NIDDK NIH HHS/United States
- R01 DK134652/DK/NIDDK NIH HHS/United States
- R01 DK067912/DK/NIDDK NIH HHS/United States
- PG/14/70/31039/BHF_/British Heart Foundation/United Kingdom
- PG/12/76/29852/BHF_/British Heart Foundation/United Kingdom
- PG/17/31/32988/BHF_/British Heart Foundation/United Kingdom
- P30 CA033572/CA/NCI NIH HHS/United States
- PG/09/052/27833/BHF_/British Heart Foundation/United Kingdom
- R01 DK112917/DK/NIDDK NIH HHS/United States
- PG/19/53/34499/BHF_/British Heart Foundation/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
