

Lack of nutritional immunity in diabetic skin infections promotes Staphylococcus aureus virulence
- PMID: 33188027
- PMCID: PMC7673755
- DOI: 10.1126/sciadv.abc5569
Lack of nutritional immunity in diabetic skin infections promotes Staphylococcus aureus virulence
Abstract
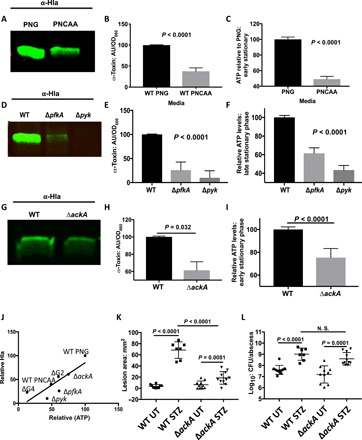
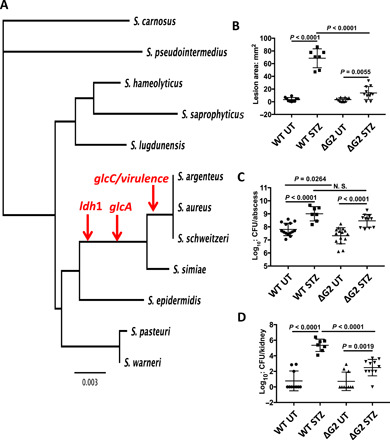
Elevated blood/tissue glucose is a hallmark feature of advanced diabetes, and people with diabetes are prone to more frequent and invasive infections with Staphylococcus aureus. Phagocytes must markedly increase glucose consumption during infection to generate and oxidative burst and kill invading bacteria. Similarly, glucose is essential for S. aureus survival in an infection and competition with the host, for this limited resource is reminiscent of nutritional immunity. Here, we show that infiltrating phagocytes do not express their high-efficiency glucose transporters in modeled diabetic infections, resulting in a diminished respiratory burst and increased glucose availability for S. aureus We show that excess glucose in these hyperglycemic abscesses significantly enhances S. aureus virulence potential, resulting in worse infection outcomes. Last, we show that two glucose transporters recently acquired by S. aureus are essential for excess virulence factor production and the concomitant increase in disease severity in hyperglycemic infections.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures




References
-
- Lipsky B. A., Tabak Y. P., Johannes R. S., Vo L., Hyde L., Weigelt J. A., Skin and soft tissue infections in hospitalised patients with diabetes: Culture isolates and risk factors associated with mortality, length of stay and cost. Diabetologia 53, 914–923 (2010). - PubMed
-
- Smit J., Søgaard M., Schønheyder H. C., Nielsen H., Frøslev T., Thomsen R. W., Diabetes and risk of community-acquired Staphylococcus aureus bacteremia: A population-based case-control study. Eur. J. Endocrinol. 174, 631–639 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
