

A Novel Role of SLC26A3 in the Maintenance of Intestinal Epithelial Barrier Integrity
- PMID: 33189700
- PMCID: PMC7956241
- DOI: 10.1053/j.gastro.2020.11.008
A Novel Role of SLC26A3 in the Maintenance of Intestinal Epithelial Barrier Integrity
Abstract
Background & aims: The down-regulated in adenoma (DRA) protein, encoded by SLC26A3, a key intestinal chloride anion exchanger, has recently been identified as a novel susceptibility gene for inflammatory bowel disease (IBD). However, the mechanisms underlying the increased susceptibility to inflammation induced by the loss of DRA remain elusive. Compromised barrier is a key event in IBD pathogenesis. The current studies were undertaken to elucidate the impact of DRA deficiency on epithelial barrier integrity and to define underlying mechanisms.
Methods: Wild-type and DRA-knockout (KO) mice and crypt-derived colonoids were used as models for intestinal epithelial response. Paracellular permeability was measured by using fluorescein isothiocyanate-dextran flux. Immunoblotting, immunofluorescence, immunohistochemistry, and ribonucleoprotein immunoprecipitation assays were performed. Gut microbiome analysis was conducted to investigate the impact of DRA deficiency on gut microbial communities.
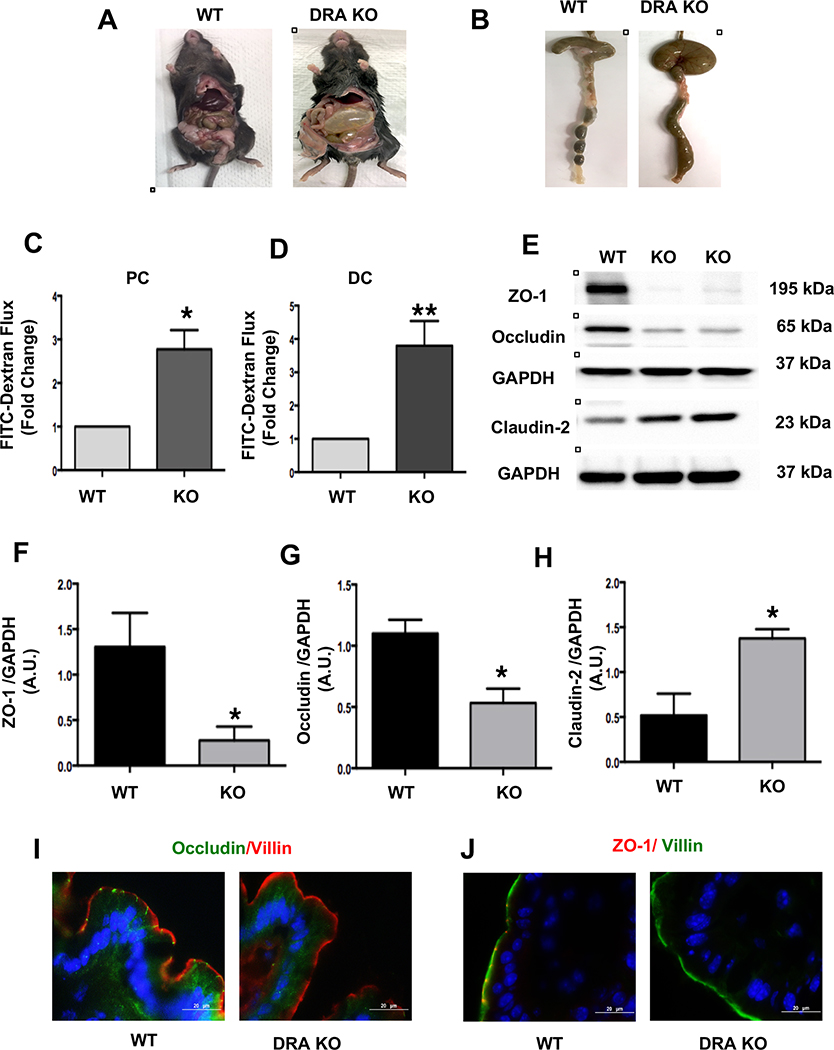
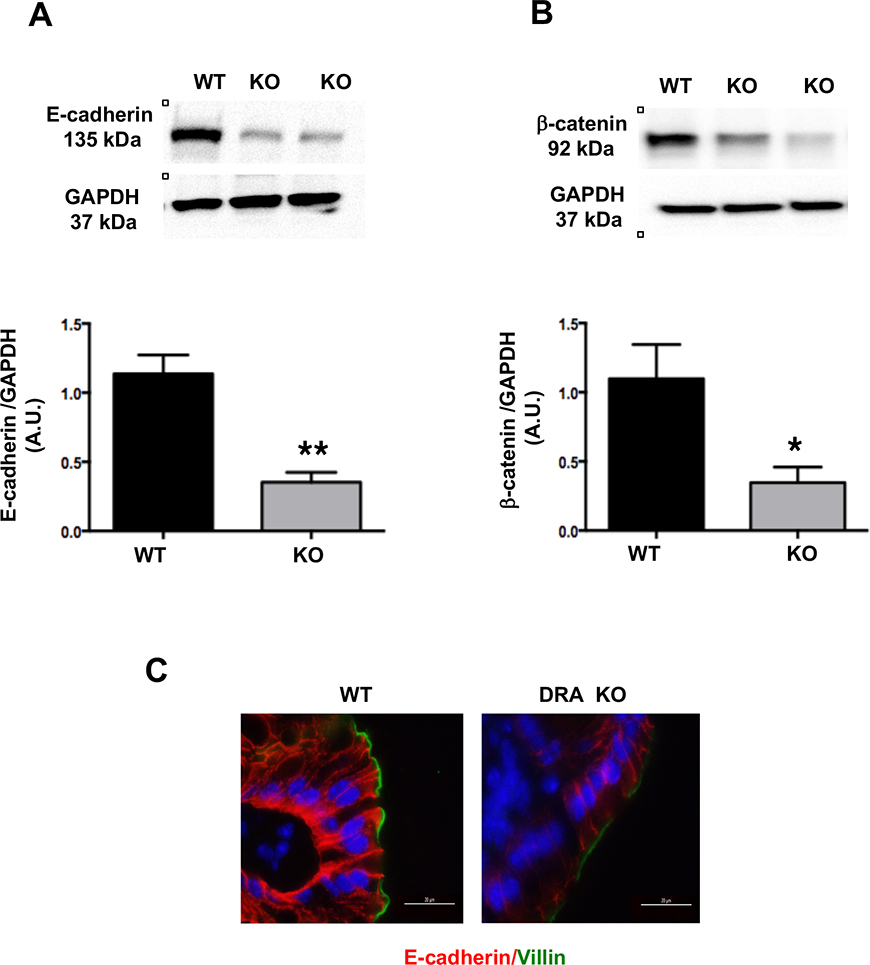
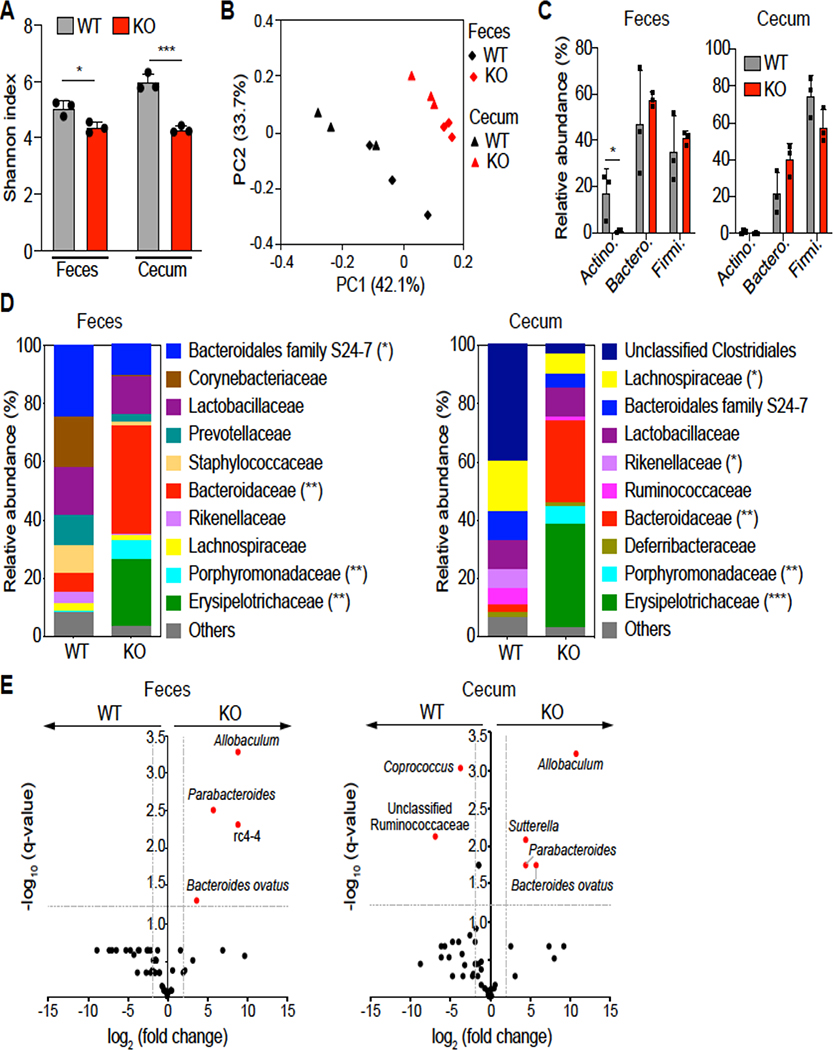
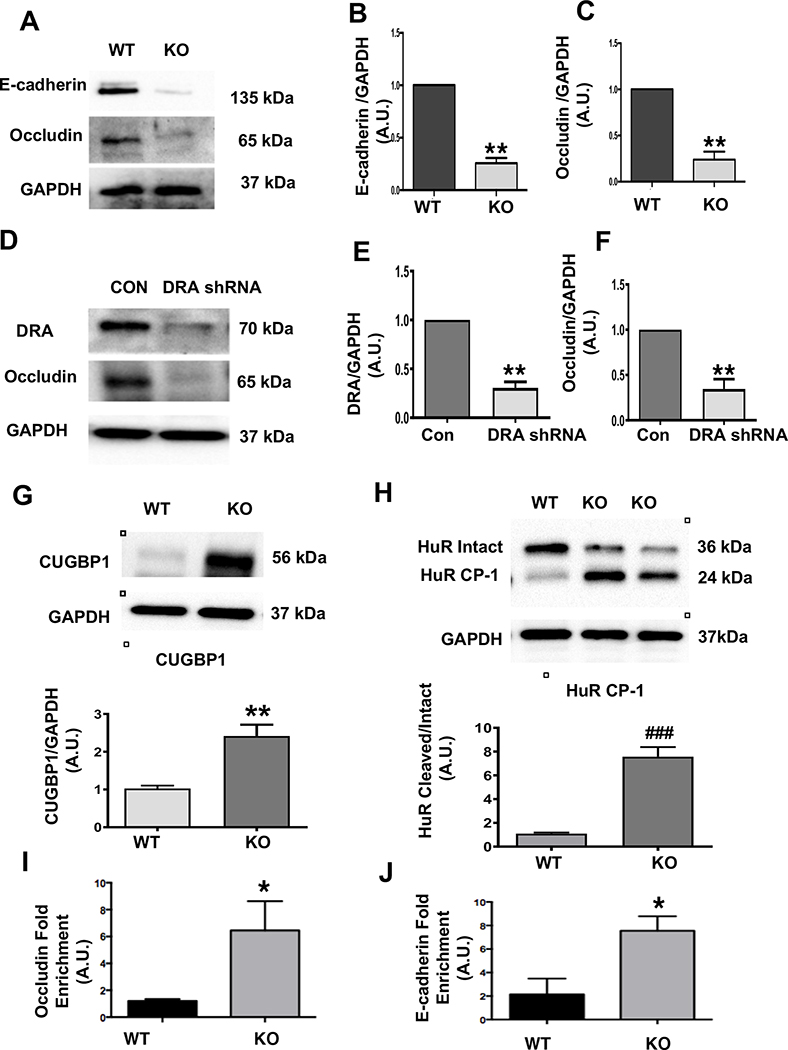
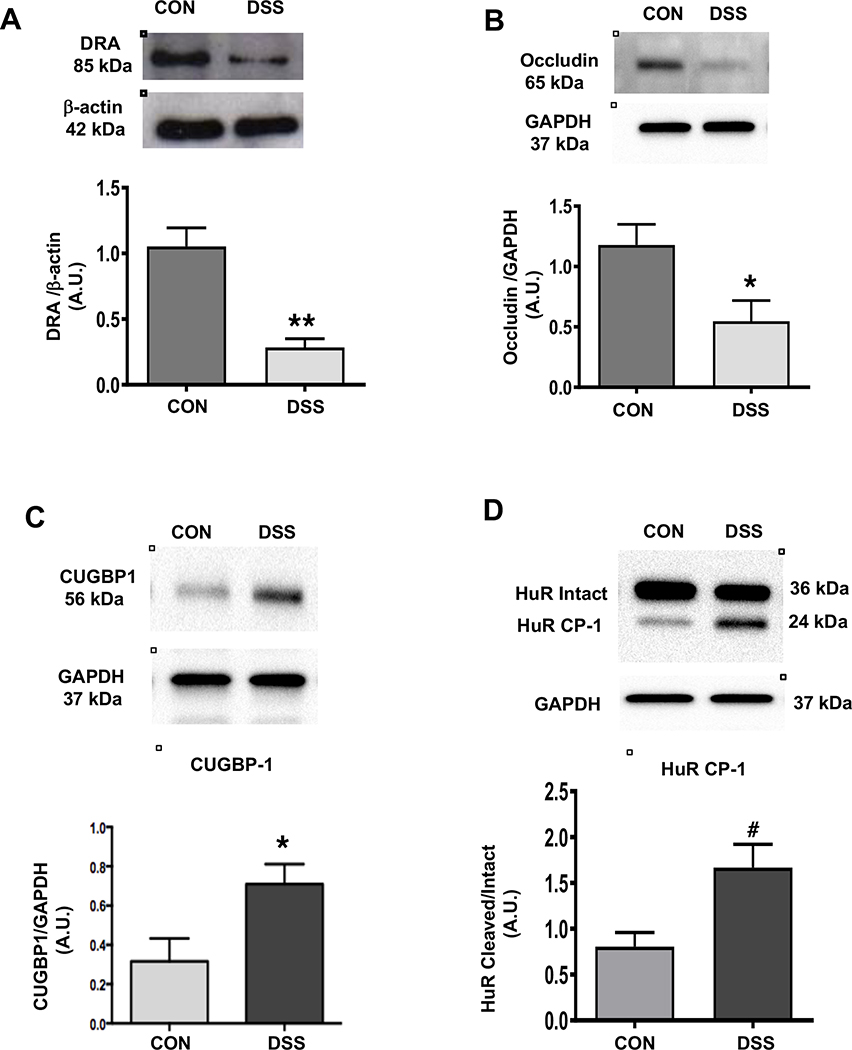
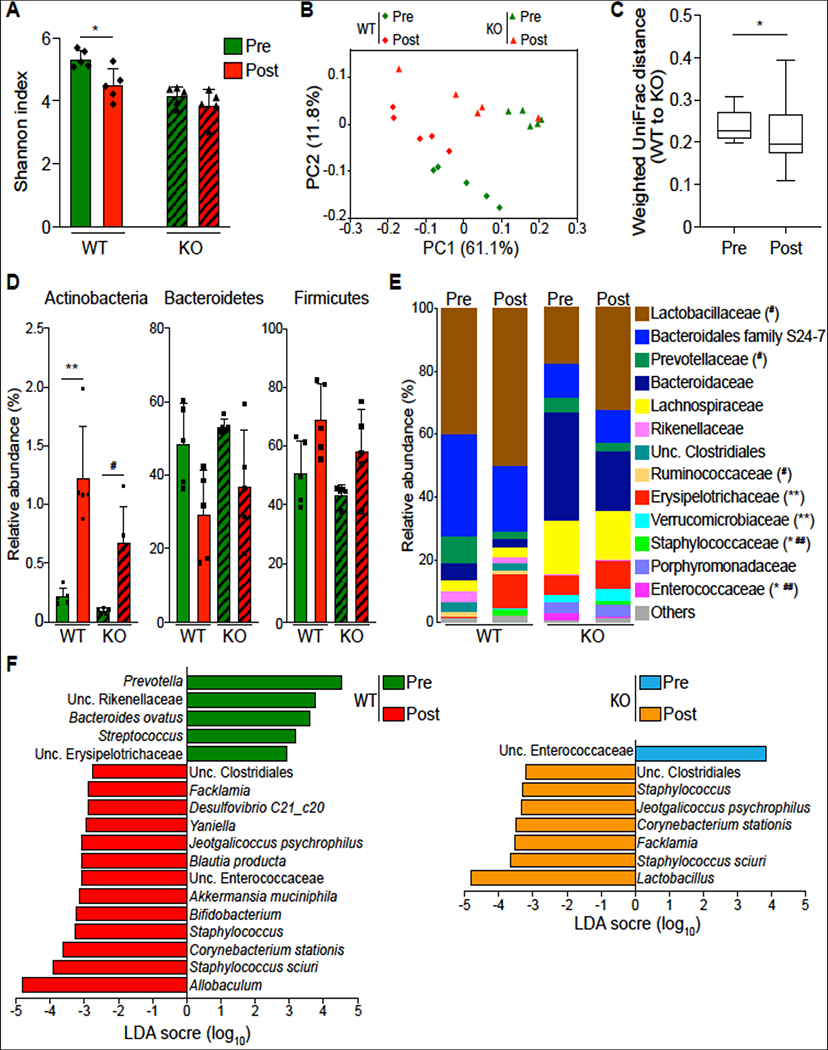
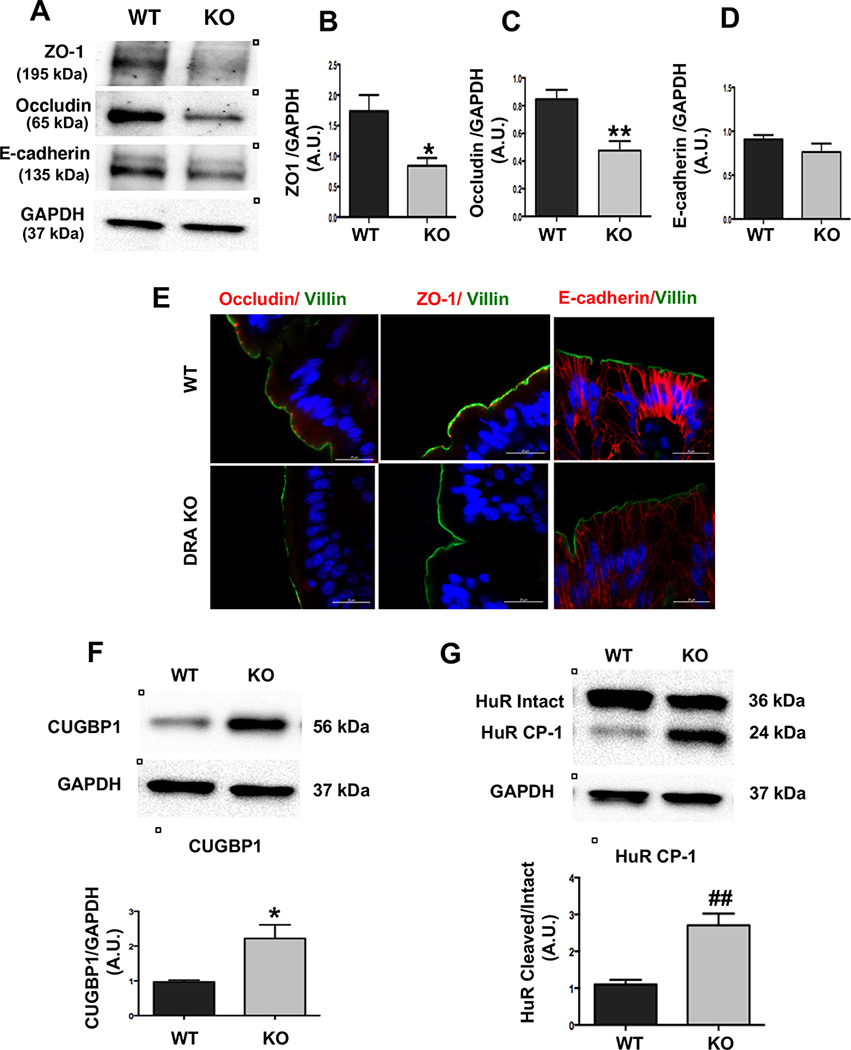
Results: DRA-KO mice exhibited an increased colonic paracellular permeability with significantly decreased levels of tight junction/adherens junction proteins, including ZO-1, occludin, and E-cadherin. A similar expression pattern of occludin and E-cadherin was observed in colonoids derived from DRA-KO mice and short hairpin RNA-mediated DRA knockdown in Caco-2 cells. Microbial analysis showed gut dysbiosis in DRA-KO mice. However, cohousing studies showed that dysbiosis played only a partial role in maintaining tight junction protein expression. Furthermore, our results showed increased binding of RNA-binding protein CUGBP1 with occludin and E-cadherin genes in DRA-KO mouse colon, suggesting that posttranscriptional mechanisms play a key role in gut barrier dysfunction.
Conclusions: To our knowledge, our studies demonstrate a novel role of DRA in maintaining the intestinal epithelial barrier function and potential implications of its dysregulation in IBD pathogenesis.
Keywords: Down-regulated in Adenoma; Gut Microbiota; Inflammatory Bowel Disease; Intestinal Chloride Transporter.
Published by Elsevier Inc.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- I01 BX002867/BX/BLRD VA/United States
- R01 DK109709/DK/NIDDK NIH HHS/United States
- R01 DK092441/DK/NIDDK NIH HHS/United States
- R56 DK092441/DK/NIDDK NIH HHS/United States
- I01 BX002011/BX/BLRD VA/United States
- IK2 BX004719/BX/BLRD VA/United States
- IK6 BX005242/BX/BLRD VA/United States
- IK6 BX005243/BX/BLRD VA/United States
- R01 DK109560/DK/NIDDK NIH HHS/United States
- R01 DK054016/DK/NIDDK NIH HHS/United States
- R01 DK098170/DK/NIDDK NIH HHS/United States
- I01 BX006177/BX/BLRD VA/United States
- I01 BX000152/BX/BLRD VA/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
