

Overlapping mechanisms of lncRNA and expanded microsatellite RNA
- PMID: 33191580
- PMCID: PMC7880542
- DOI: 10.1002/wrna.1634
Overlapping mechanisms of lncRNA and expanded microsatellite RNA
Abstract
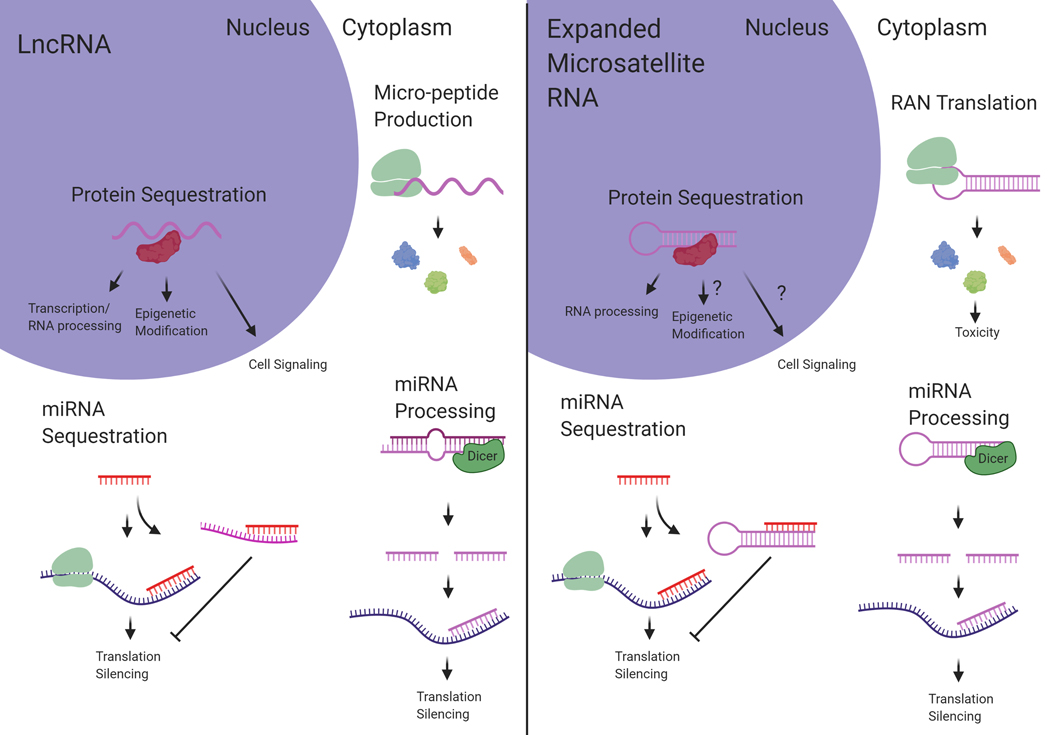
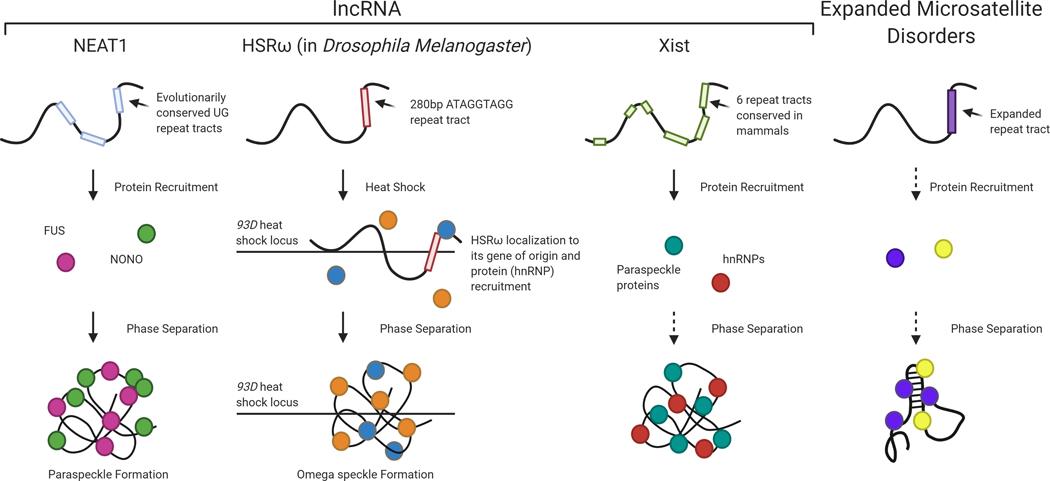
RNA has major regulatory roles in a wide range of biological processes and a surge of RNA research has led to the classification of numerous functional RNA species. One example is long noncoding RNAs (lncRNAs) that are structurally complex transcripts >200 nucleotides (nt) in length and lacking a canonical open reading frame (ORF). Despite a general lack of sequence conservation and low expression levels, many lncRNAs have been shown to have functionality in diverse biological processes as well as in mechanisms of disease. In parallel with the growing understanding of lncRNA functions, there is a growing subset of microsatellite expansion disorders in which the primary mechanism of pathogenesis is an RNA gain of function arising from RNA transcripts from the mutant allele. Microsatellite expansion disorders are caused by an expansion of short (3-10 nt) repeats located within coding genes. Expanded repeat-containing RNA mediates toxicity through multiple mechanisms, the details of which remain only partially understood. The purpose of this review is to highlight the links between functional mechanisms of lncRNAs and the potential pathogenic mechanisms of expanded microsatellite RNA. These shared mechanisms include protein sequestration, peptide translation, micro-RNA (miRNA) processing, and miRNA sequestration. Recognizing the parallels between the normal functions of lncRNAs and the negative impact of expanded microsatellite RNA on biological processes can provide reciprocal understanding to the roles of both RNA species. This article is categorized under: RNA Interactions with Proteins and Other Molecules > Protein-RNA Interactions: Functional Implications RNA in Disease and Development > RNA in Disease.
Keywords: expanded microsatellite RNA; long noncoding RNA; miRNA; protein sequestration.
© 2020 Wiley Periodicals LLC.
Figures
References
-
- Aly MK, Ninomiya K, Adachi S, Natsume T, & Hirose T. (2019). Two distinct nuclear stress bodies containing different sets of RNA-binding proteins are formed with HSATIII architectural noncoding RNAs upon thermal stress exposure. Biochemical and Biophysical Research Communications, 516(2), 419–423. doi: 10.1016/j.bbrc.2019.06.061 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
