

Object Recognition Memory: Distinct Yet Complementary Roles of the Mouse CA1 and Perirhinal Cortex
- PMID: 33192287
- PMCID: PMC7642692
- DOI: 10.3389/fnmol.2020.527543
Object Recognition Memory: Distinct Yet Complementary Roles of the Mouse CA1 and Perirhinal Cortex
Abstract
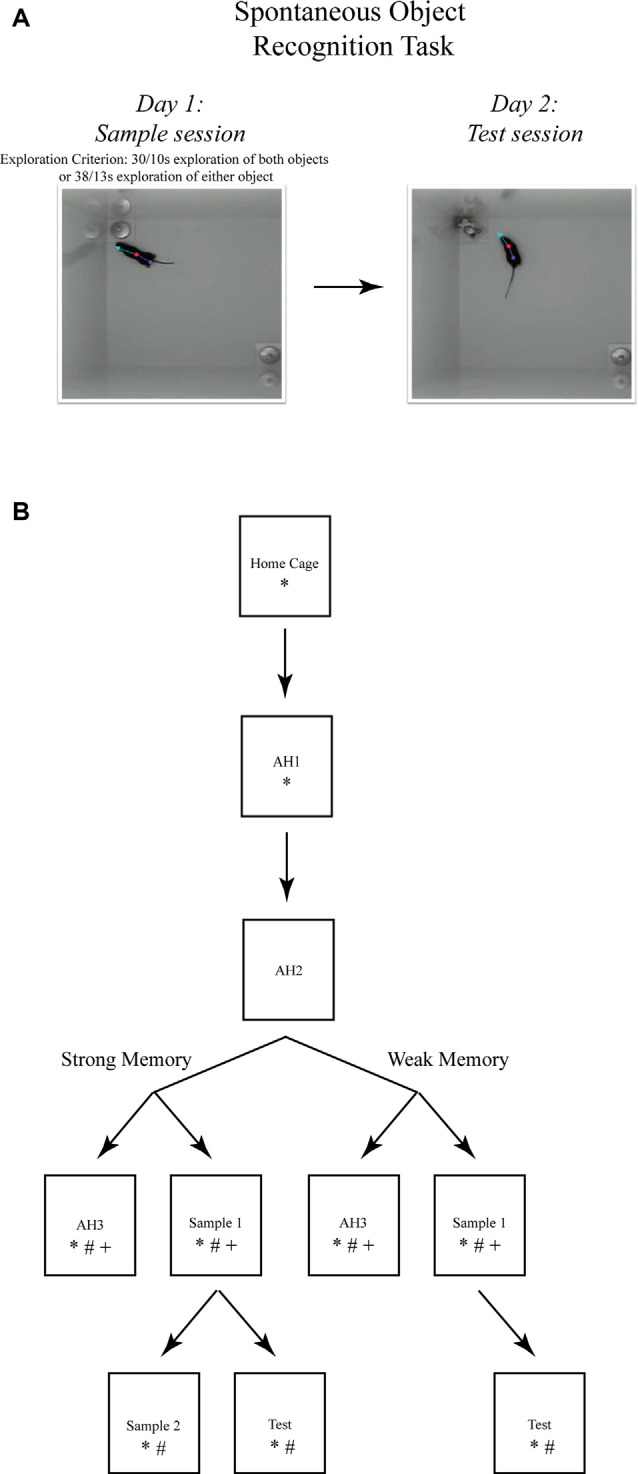
While the essential contribution of the hippocampus to spatial memory is well established, object recognition memory has been traditionally attributed to the perirhinal cortex (PRh). However, the results of several studies indicate that under specific procedural conditions, temporary or permanent lesions of the hippocampus affect object memory processes as measured in the Spontaneous Object Recognition (SOR) task. The PRh and hippocampus are considered to contribute distinctly to object recognition memory based on memory strength. Allowing mice more, or less, exploration of novel objects during the encoding phase of the task (i.e., sample session), yields stronger, or weaker, object memory, respectively. The current studies employed temporary local inactivation and immunohistochemistry to determine the differential contributions of neuronal activity in PRh and the CA1 region of the hippocampus to strong and weak object memory. Temporary inactivation of the CA1 immediately after the SOR sample session impaired strong object memory but spared weak object memory; while temporary inactivation of PRh post-sample impaired weak object memory but spared strong object memory. Furthermore, mRNA transcription and de novo protein synthesis are required for the consolidation of episodic memory, and activation patterns of immediate early genes (IEGs), such as c-Fos and Arc, are linked to behaviorally triggered neuronal activation and synaptic plasticity. Analyses of c-Fos and Arc protein expression in PRh and CA1 neurons by immunohistochemistry, and of Arc mRNA by qPCR after distinct stages of SOR, provide additional support that strong object memory is dependent on CA1 neuronal activity, while weak object memory is dependent on PRh neuronal activity. Taken together, the results support the view that both PRh and CA1 are required for object memory under distinct conditions. Specifically, our results are consistent with a model that as the mouse begins to explore a novel object, information about it accumulates within PRh, and a weak memory of the object is encoded. If object exploration continues beyond some threshold, strong memory for the event of object exploration is encoded; the consolidation of which is CA1-dependent. These data serve to reconcile the dissension in the literature by demonstrating functional and complementary roles for CA1 and PRh neurons in rodent object memory.
Keywords: Arc; hippocampus; muscimol; object recognition; qRT-PCR.
Copyright © 2020 Cinalli, Cohen, Guthrie and Stackman.
Figures



References
-
- Barbosa F. F., Santos J. R., Meurer Y. S., Macedo P. T., Ferreira L. M., Pontes I. M., et al. . (2013). Differential cortical c-Fos and Zif-268 expression after object and spatial memory processing in a standard or episodic-like object recognition task. Front. Behav. Neurosci. 7:112. 10.3389/fnbeh.2013.00112 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous