

Tick-transmitted thogotovirus gains high virulence by a single MxA escape mutation in the viral nucleoprotein
- PMID: 33196685
- PMCID: PMC7704052
- DOI: 10.1371/journal.ppat.1009038
Tick-transmitted thogotovirus gains high virulence by a single MxA escape mutation in the viral nucleoprotein
Abstract
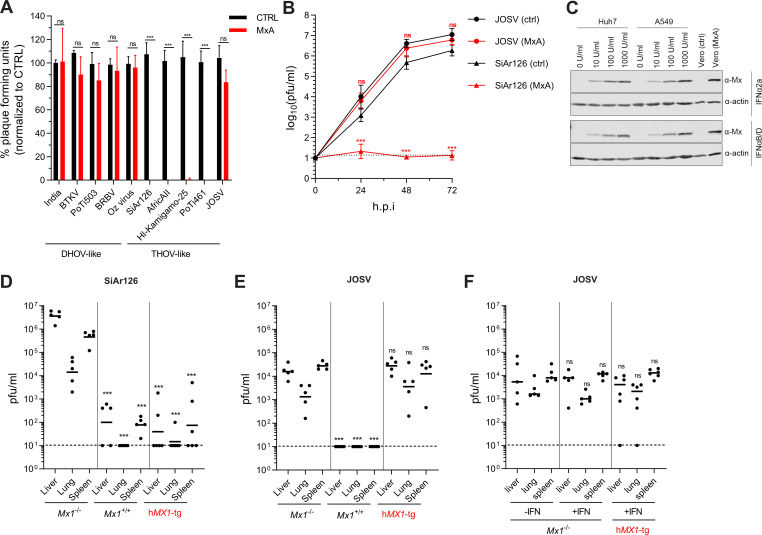
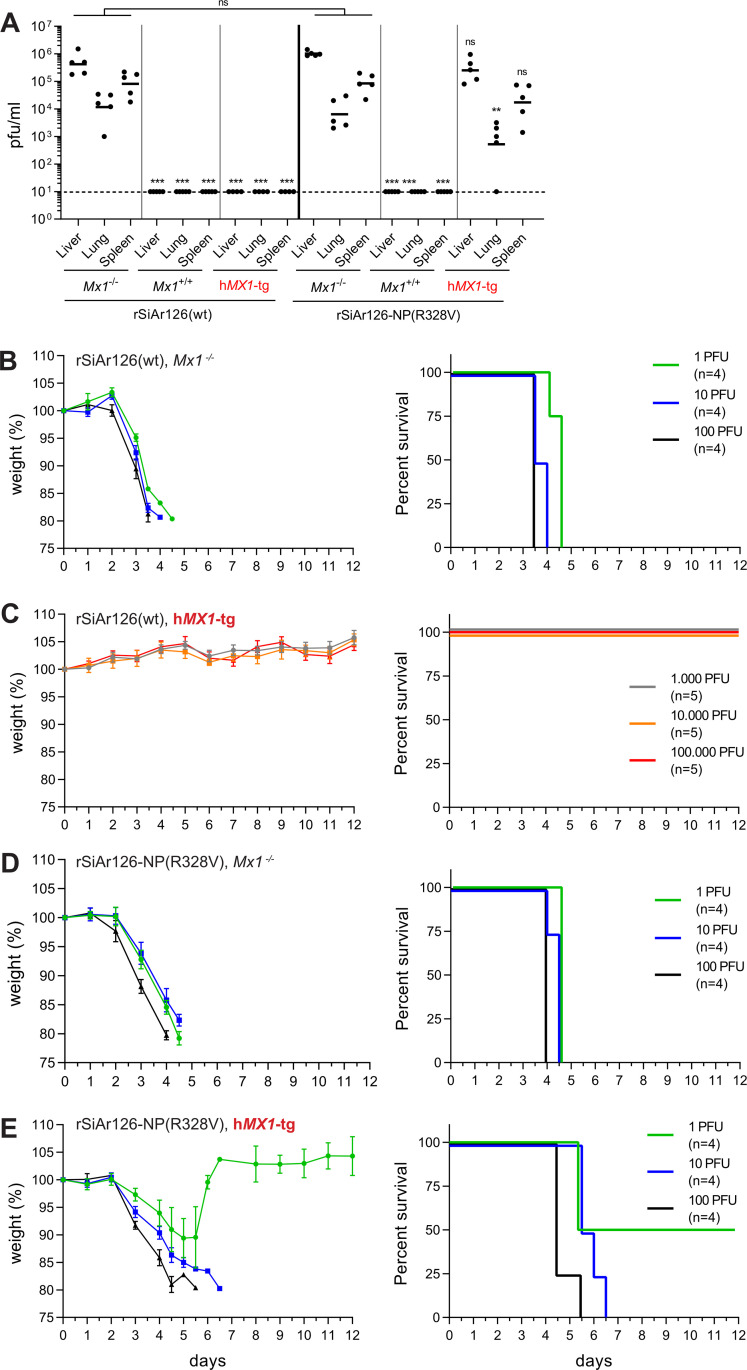
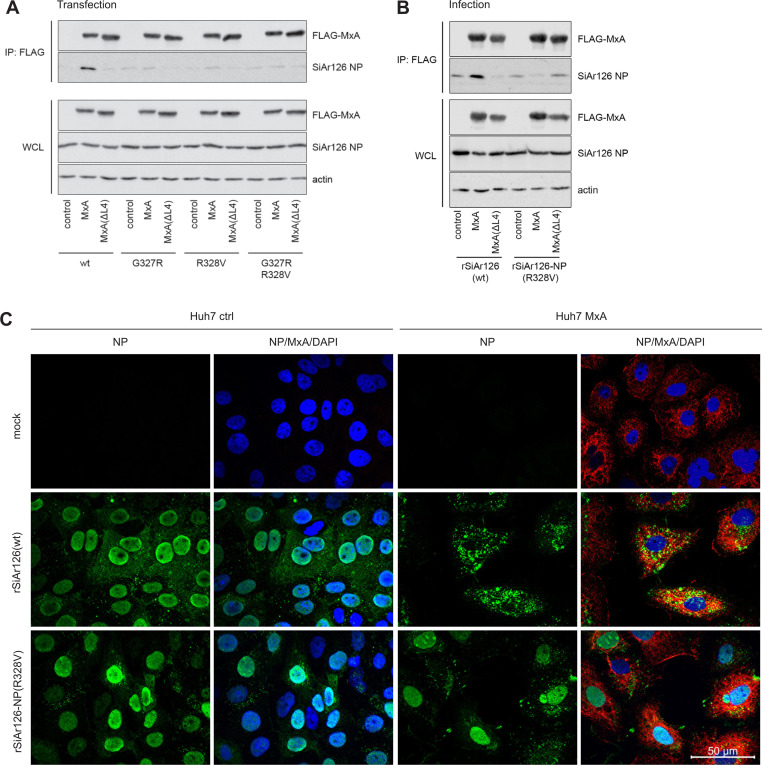
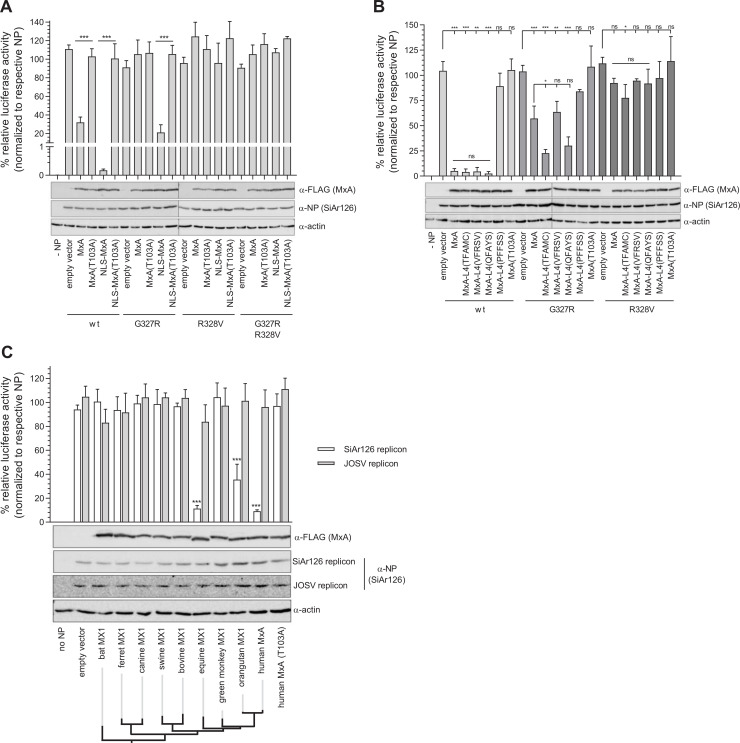
Infections with emerging and re-emerging arboviruses are of increasing concern for global health. Tick-transmitted RNA viruses of the genus Thogotovirus in the Orthomyxoviridae family have considerable zoonotic potential, as indicated by the recent emergence of Bourbon virus in the USA. To successfully infect humans, arboviruses have to escape the restrictive power of the interferon defense system. This is exemplified by the high sensitivity of thogotoviruses to the antiviral action of the interferon-induced myxovirus resistance protein A (MxA) that inhibits the polymerase activity of incoming viral ribonucleoprotein complexes. Acquiring resistance to human MxA would be expected to enhance the zoonotic potential of these pathogens. Therefore, we screened a panel of 10 different thogotovirus isolates obtained from various parts of the world for their sensitivity to MxA. A single isolate from Nigeria, Jos virus, showed resistance to the antiviral action of MxA in cell culture and in MxA-transgenic mice, whereas the prototypic Sicilian isolate SiAr126 was fully MxA-sensitive. Further analysis identified two amino acid substitutions (G327R and R328V) in the viral nucleoprotein as determinants for MxA resistance. Importantly, when introduced into SiAr126, the R328V mutation resulted in complete MxA escape of the recombinant virus, without causing any viral fitness loss. The escape mutation abolished viral nucleoprotein recognition by MxA and allowed unhindered viral growth in MxA-expressing cells and in MxA-transgenic mice. These findings demonstrate that thogotoviruses can overcome the species barrier by escaping MxA restriction and reveal that these tick-transmitted viruses may have a greater zoonotic potential than previously suspected.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Eurasian Avian-Like Swine Influenza A Viruses Escape Human MxA Restriction through Distinct Mutations in Their Nucleoprotein.J Virol. 2019 Jan 4;93(2):e00997-18. doi: 10.1128/JVI.00997-18. J Virol. 2019. PMID: 30355693 Free PMC article.
-
Comparative Study of Ten Thogotovirus Isolates and Their Distinct In Vivo Characteristics.J Virol. 2022 Mar 9;96(5):e0155621. doi: 10.1128/JVI.01556-21. Epub 2022 Jan 12. J Virol. 2022. PMID: 35019718 Free PMC article.
-
Combinatorial mutagenesis of rapidly evolving residues yields super-restrictor antiviral proteins.PLoS Biol. 2019 Oct 1;17(10):e3000181. doi: 10.1371/journal.pbio.3000181. eCollection 2019 Oct. PLoS Biol. 2019. PMID: 31574080 Free PMC article.
-
Mx genes: host determinants controlling influenza virus infection and trans-species transmission.Hum Genet. 2020 Jun;139(6-7):695-705. doi: 10.1007/s00439-019-02092-8. Epub 2019 Nov 26. Hum Genet. 2020. PMID: 31773252 Free PMC article. Review.
-
[Research advances on thogoto virus of the family orthomyxoviridae].Bing Du Xue Bao. 2013 Sep;29(5):555-8. Bing Du Xue Bao. 2013. PMID: 24386846 Review. Chinese.
Cited by
-
Heterozygous and generalist MxA super-restrictors overcome breadth-specificity tradeoffs in antiviral restriction.bioRxiv [Preprint]. 2024 Oct 10:2024.10.10.617484. doi: 10.1101/2024.10.10.617484. bioRxiv. 2024. Update in: Sci Adv. 2025 May 2;11(18):eadu0062. doi: 10.1126/sciadv.adu0062. PMID: 39416221 Free PMC article. Updated. Preprint.
-
Comprehensive Review of Emergence and Virology of Tickborne Bourbon Virus in the United States.Emerg Infect Dis. 2023 Jan;29(1):1-7. doi: 10.3201/eid2901.212295. Emerg Infect Dis. 2023. PMID: 36573641 Free PMC article. Review.
-
Cryo-EM structures of Thogoto virus polymerase reveal unique RNA transcription and replication mechanisms among orthomyxoviruses.Nat Commun. 2024 May 30;15(1):4620. doi: 10.1038/s41467-024-48848-3. Nat Commun. 2024. PMID: 38816392 Free PMC article.
-
The Antiviral Activity of Equine Mx1 against Thogoto Virus Is Determined by the Molecular Structure of Its Viral Specificity Region.J Virol. 2023 Feb 28;97(2):e0193822. doi: 10.1128/jvi.01938-22. Epub 2023 Feb 7. J Virol. 2023. PMID: 36749070 Free PMC article.
-
Heterozygous and generalist MxA super-restrictors overcome breadth-specificity trade-offs in antiviral restriction.Sci Adv. 2025 May 2;11(18):eadu0062. doi: 10.1126/sciadv.adu0062. Epub 2025 May 2. Sci Adv. 2025. PMID: 40315333 Free PMC article.
References
-
- McCauley JW, Hongo S, Kaverin N V, Kochs G, Lamb RA, Matrosovich MN, et al. Family orthomyxoviridae. Virus Taxon ninth Rep Int Comm Taxon viruses. 2012;749–761.
-
- Anderson CR, Casals J. Dhori virus, a new agent isolated from Hyalomma dromedarii in India. Indian J Med Res. 1973. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
