

Antipsychotic olanzapine-induced misfolding of proinsulin in the endoplasmic reticulum accounts for atypical development of diabetes
- PMID: 33198886
- PMCID: PMC7671685
- DOI: 10.7554/eLife.60970
Antipsychotic olanzapine-induced misfolding of proinsulin in the endoplasmic reticulum accounts for atypical development of diabetes
Abstract
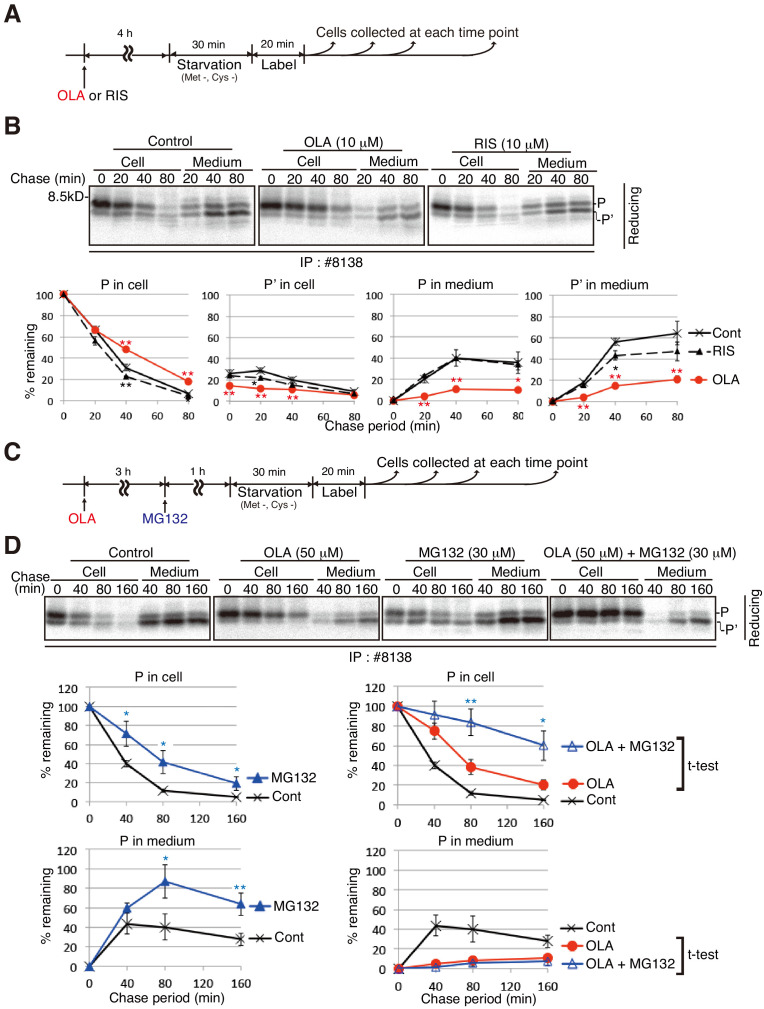
Second-generation antipsychotics are widely used to medicate patients with schizophrenia, but may cause metabolic side effects such as diabetes, which has been considered to result from obesity-associated insulin resistance. Olanzapine is particularly well known for this effect. However, clinical studies have suggested that olanzapine-induced hyperglycemia in certain patients cannot be explained by such a generalized mechanism. Here, we focused on the effects of olanzapine on insulin biosynthesis and secretion by mouse insulinoma MIN6 cells. Olanzapine reduced maturation of proinsulin, and thereby inhibited secretion of insulin; and specifically shifted the primary localization of proinsulin from insulin granules to the endoplasmic reticulum. This was due to olanzapine's impairment of proper disulfide bond formation in proinsulin, although direct targets of olanzapine remain undetermined. Olanzapine-induced proinsulin misfolding and subsequent decrease also occurred at the mouse level. This mechanism of olanzapine-induced β-cell dysfunction should be considered, together with weight gain, when patients are administered olanzapine.
Keywords: cell biology; degradation; diabetes; misfolding; mouse; proinsulin.
© 2020, Ninagawa et al.
Conflict of interest statement
SN, ST, MO, KI, MK, SK, KI, HU, TI, RM, ST, YI, KI, TA, TN, KM No competing interests declared
Figures










References
-
- Atkins C, Liu Q, Minthorn E, Zhang SY, Figueroa DJ, Moss K, Stanley TB, Sanders B, Goetz A, Gaul N, Choudhry AE, Alsaid H, Jucker BM, Axten JM, Kumar R. Characterization of a novel PERK kinase inhibitor with antitumor and antiangiogenic activity. Cancer Research. 2013;73:1993–2002. doi: 10.1158/0008-5472.CAN-12-3109. - DOI - PubMed
-
- Carrillo JA, Herráiz AG, Ramos SI, Gervasini G, Vizcaíno S, Benítez J. Role of the Smoking-Induced cytochrome P450 (CYP)1A2 and polymorphic CYP2D6 in Steady-State concentration of olanzapine. Journal of Clinical Psychopharmacology. 2003;23:119–127. doi: 10.1097/00004714-200304000-00003. - DOI - PubMed
-
- Chang SG, Choi KD, Jang SH, Shin HC. Role of disulfide bonds in the structure and activity of human insulin. Molecules and Cells. 2003;16:323–330. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 18K06216/Ministry of Education, Culture, Sports, Science and Technology/International
- 19K06658/Ministry of Education, Culture, Sports, Science and Technology/International
- 17H01432/Ministry of Education, Culture, Sports, Science and Technology/International
- 17H06419/Ministry of Education, Culture, Sports, Science and Technology/International
