

Metaplasticity in the Ventral Pallidum as a Potential Marker for the Propensity to Gain Weight in Chronic High-Calorie Diet
- PMID: 33199503
- PMCID: PMC7726535
- DOI: 10.1523/JNEUROSCI.1809-20.2020
Metaplasticity in the Ventral Pallidum as a Potential Marker for the Propensity to Gain Weight in Chronic High-Calorie Diet
Abstract
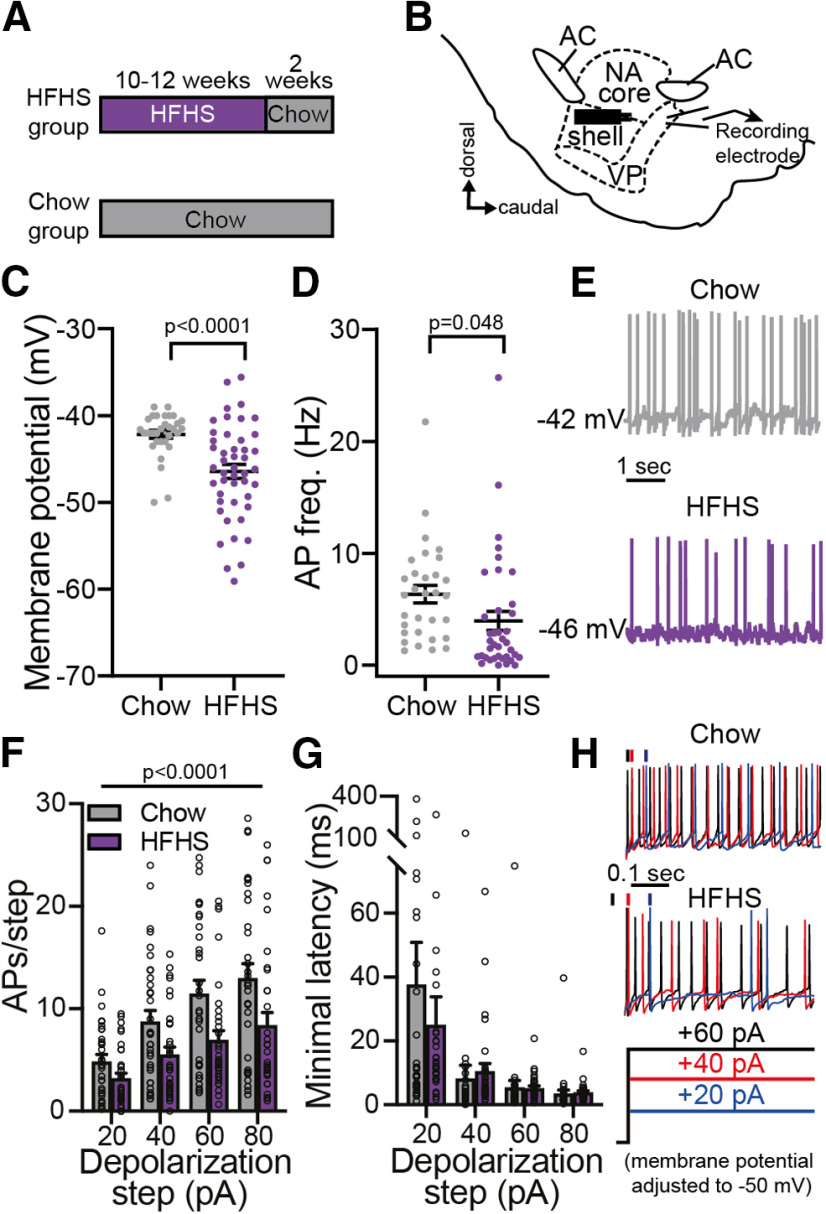
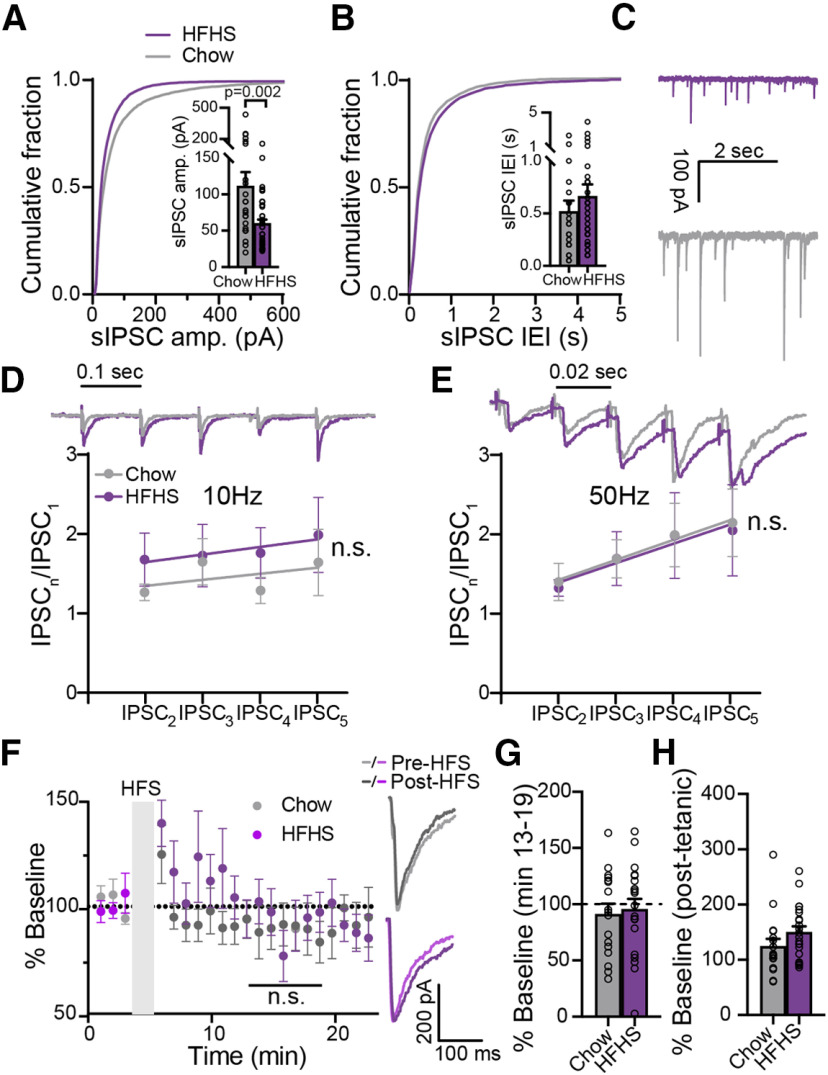
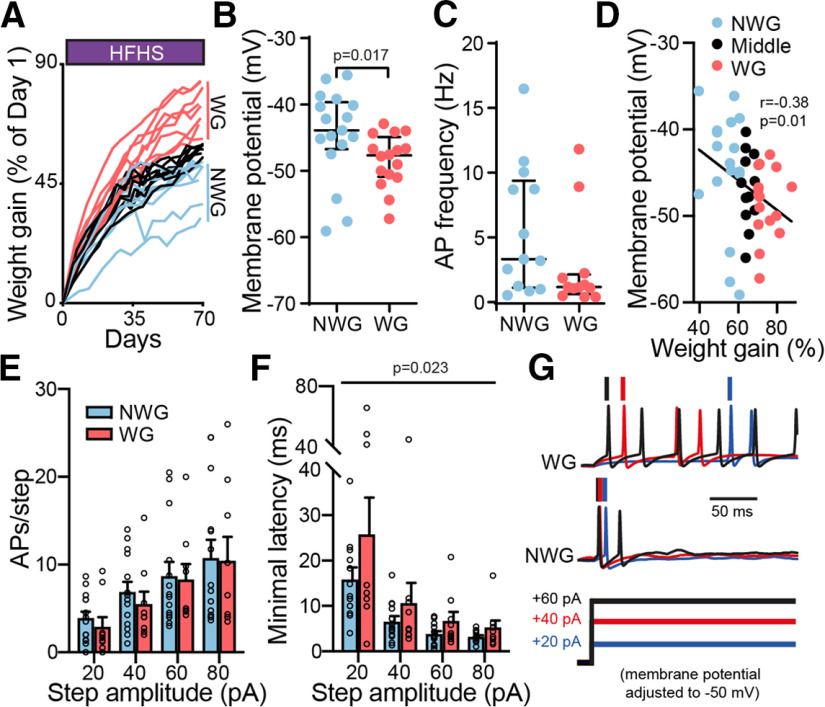
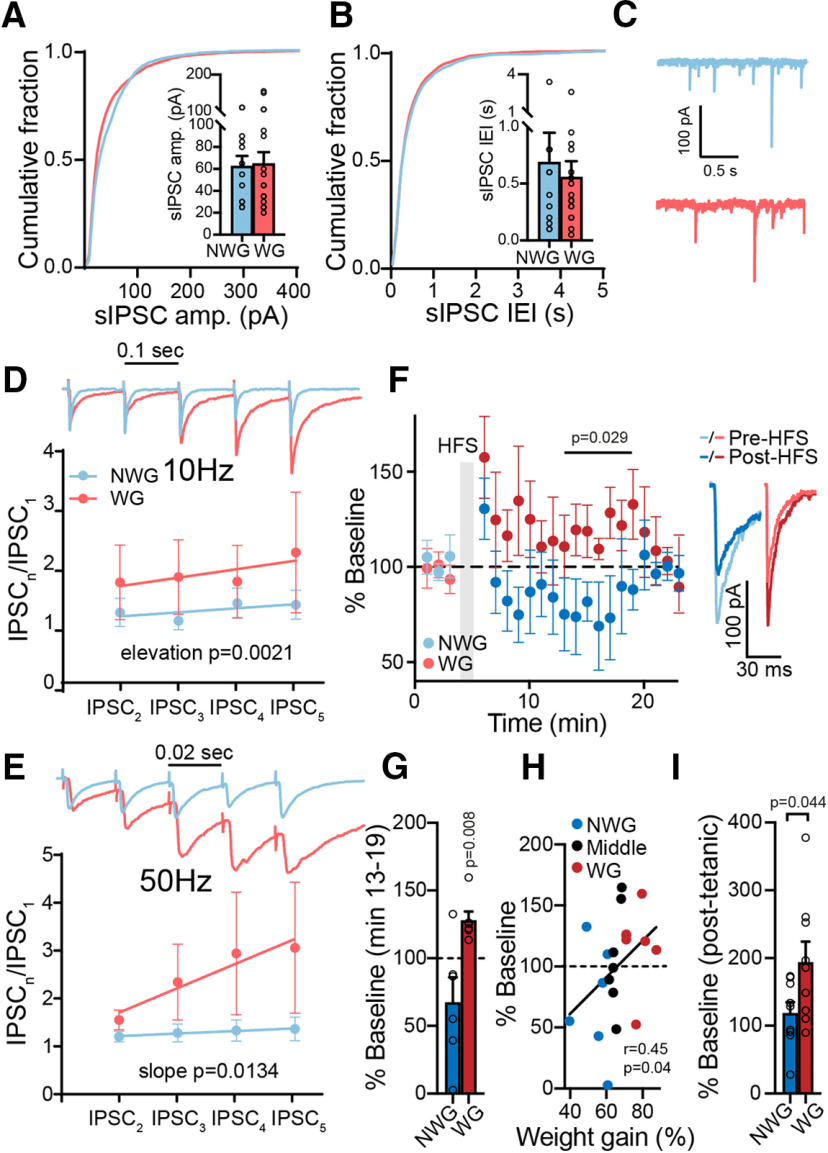
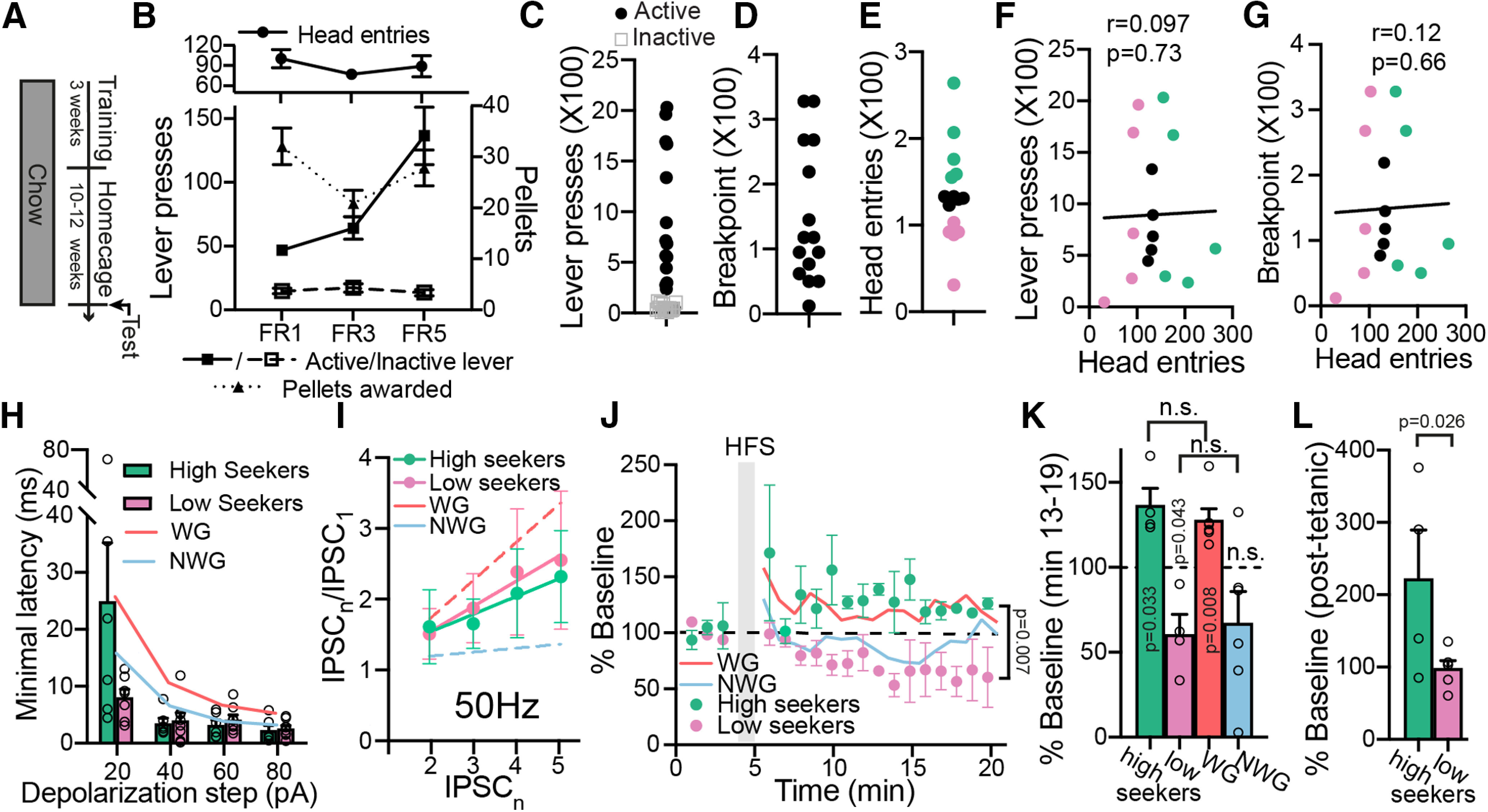
A major driver of obesity is the increasing palatability of processed foods. Although reward circuits promote the consumption of palatable food, their involvement in obesity remains unclear. The ventral pallidum (VP) is a key hub in the reward system that encodes the hedonic aspects of palatable food consumption and participates in various proposed feeding circuits. However, there is still no evidence for its involvement in developing diet-induced obesity. Here we examine, using male C57BL6/J mice and patch-clamp electrophysiology, how chronic high-fat high-sugar (HFHS) diet changes the physiology of the VP and whether mice that gain the most weight differ in their VP physiology from others. We found that 10-12 weeks of HFHS diet hyperpolarized and decreased the firing rate of VP neurons without a major change in synaptic inhibitory input. Within the HFHS group, the top 33% weight gainers (WGs) had a more hyperpolarized VP with longer latency to fire action potentials on depolarization compared with bottom 33% of weight gainers (i.e., non-weight gainers). WGs also showed synaptic potentiation of inhibitory inputs both at the millisecond and minute ranges. Moreover, we found that the tendency to potentiate the inhibitory inputs to the VP might exist in overeating mice even before exposure to HFHS, thus making it a potential property of being an overeater. These data point to the VP as a critical player in obesity and suggest that hyperpolarized membrane potential of, and potentiated inhibitory inputs to, VP neurons may play a significant role in promoting the overeating of palatable food.SIGNIFICANCE STATEMENT In modern world, where highly palatable food is readily available, overeating is often driven by motivational, rather than metabolic, needs. It is thus conceivable that reward circuits differ between obese and normal-weight individuals. But is such difference, if it exists, innate or does it develop with overeating? Here we reveal synaptic properties in the ventral pallidum, a central hub of reward circuits, that differ between mice that gain the most and the least weight when given unlimited access to highly palatable food. We show that these synaptic differences also exist without exposure to palatable food, potentially making them innate properties that render some more susceptible than others to overeat. Thus, the propensity to overeat may have a strong innate component embedded in reward circuits.
Keywords: motivation; obesity; overeating; patch clamp; synaptic plasticity; ventral pallidum.
Copyright © 2020 the authors.
Figures
Similar articles
-
Dopamine D1 receptor gene expression decreases in the nucleus accumbens upon long-term exposure to palatable food and differs depending on diet-induced obesity phenotype in rats.Neuroscience. 2010 Dec 15;171(3):779-87. doi: 10.1016/j.neuroscience.2010.09.046. Epub 2010 Sep 26. Neuroscience. 2010. PMID: 20875839
-
Neural Activity in the Ventral Pallidum Encodes Variation in the Incentive Value of a Reward Cue.J Neurosci. 2016 Jul 27;36(30):7957-70. doi: 10.1523/JNEUROSCI.0736-16.2016. J Neurosci. 2016. PMID: 27466340 Free PMC article.
-
Projection-Specific Potentiation of Ventral Pallidal Glutamatergic Outputs after Abstinence from Cocaine.J Neurosci. 2020 Feb 5;40(6):1276-1285. doi: 10.1523/JNEUROSCI.0929-19.2019. Epub 2019 Dec 13. J Neurosci. 2020. PMID: 31836662 Free PMC article.
-
The role of the nucleus accumbens and ventral pallidum in feeding and obesity.Prog Neuropsychopharmacol Biol Psychiatry. 2021 Dec 20;111:110394. doi: 10.1016/j.pnpbp.2021.110394. Epub 2021 Jul 6. Prog Neuropsychopharmacol Biol Psychiatry. 2021. PMID: 34242717 Review.
-
Ventral pallidal regulation of motivated behaviors and reinforcement.Front Neural Circuits. 2023 Feb 2;17:1086053. doi: 10.3389/fncir.2023.1086053. eCollection 2023. Front Neural Circuits. 2023. PMID: 36817646 Free PMC article. Review.
Cited by
-
Ventral Pallidal GABAergic Neurons Drive Consumption in Male, But Not Female, Rats.eNeuro. 2025 Feb 3;12(2):ENEURO.0245-24.2025. doi: 10.1523/ENEURO.0245-24.2025. Print 2025 Feb. eNeuro. 2025. PMID: 39809537 Free PMC article.
-
Transcriptomic landscape of mammalian ventral pallidum at single-cell resolution.Sci Adv. 2024 Dec 13;10(50):eadq6017. doi: 10.1126/sciadv.adq6017. Epub 2024 Dec 11. Sci Adv. 2024. PMID: 39661664 Free PMC article.
-
Chronic chemogenetic manipulation of ventral pallidum targeted neurons in male rats fed an obesogenic diet.Brain Res. 2022 Jun 1;1784:147886. doi: 10.1016/j.brainres.2022.147886. Epub 2022 Mar 18. Brain Res. 2022. PMID: 35314149 Free PMC article.
-
Functional Magnetic Resonance Imaging and Obesity-Novel Ways to Seen the Unseen.J Clin Med. 2022 Jun 20;11(12):3561. doi: 10.3390/jcm11123561. J Clin Med. 2022. PMID: 35743630 Free PMC article. Review.
-
Multimodal Interrogation of Ventral Pallidum Projections Reveals Projection-Specific Signatures and Effects on Cocaine Reward.J Neurosci. 2024 May 1;44(18):e1469232024. doi: 10.1523/JNEUROSCI.1469-23.2024. J Neurosci. 2024. PMID: 38485256 Free PMC article.
References
-
- Alonso-Caraballo Y, Fetterly TL, Jorgensen ET, Nieto AM, Brown TE, Ferrario CR (2020) Sex specific effects of “junk-food” diet on calcium permeable AMPA receptors and silent synapses in the nucleus accumbens core. Neuropsychopharmacology. Advance online publication. Retrieved July 20, 2020. doi: 10.1038/s41386-020-0781-1 10.1038/s41386-020-0781-1 - DOI - DOI - PMC - PubMed
-
- Alsiö J, Olszewski PK, Norbäck AH, Gunnarsson ZEA, Levine AS, Pickering C, Schiöth HB (2010) Dopamine D1 receptor gene expression decreases in the nucleus accumbens upon long-term exposure to palatable food and differs depending on diet-induced obesity phenotype in rats. Neuroscience 171:779–787. 10.1016/j.neuroscience.2010.09.046 - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources