

Structure of the helicase core of Werner helicase, a key target in microsatellite instability cancers
- PMID: 33199508
- PMCID: PMC7671478
- DOI: 10.26508/lsa.202000795
Structure of the helicase core of Werner helicase, a key target in microsatellite instability cancers
Abstract
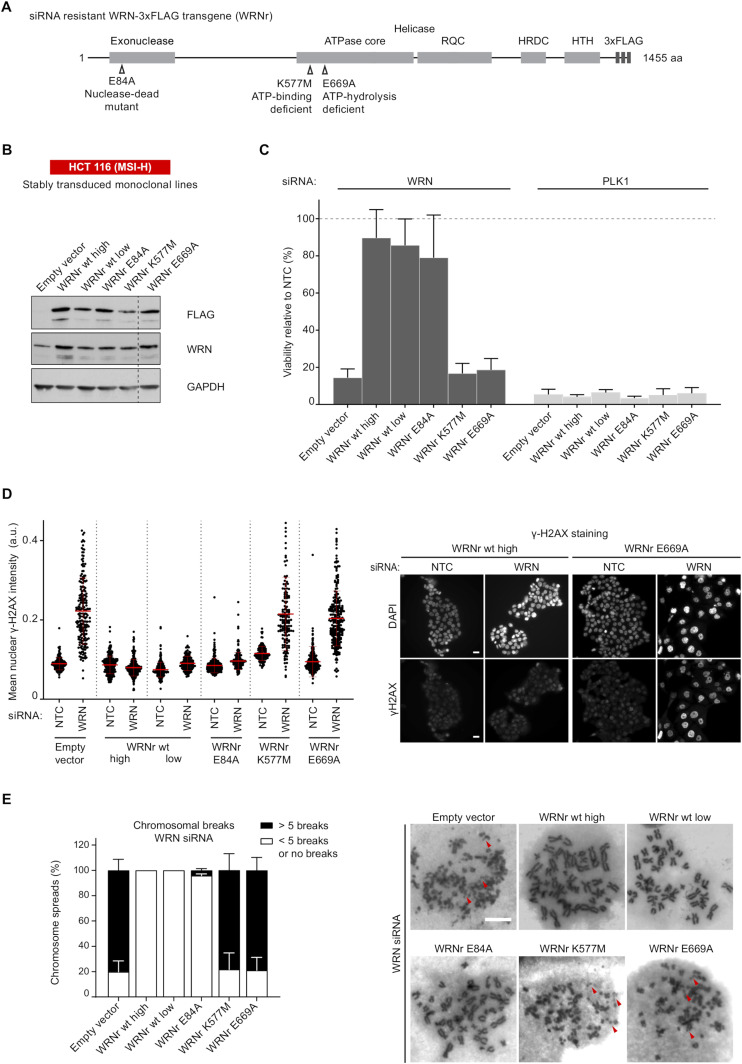
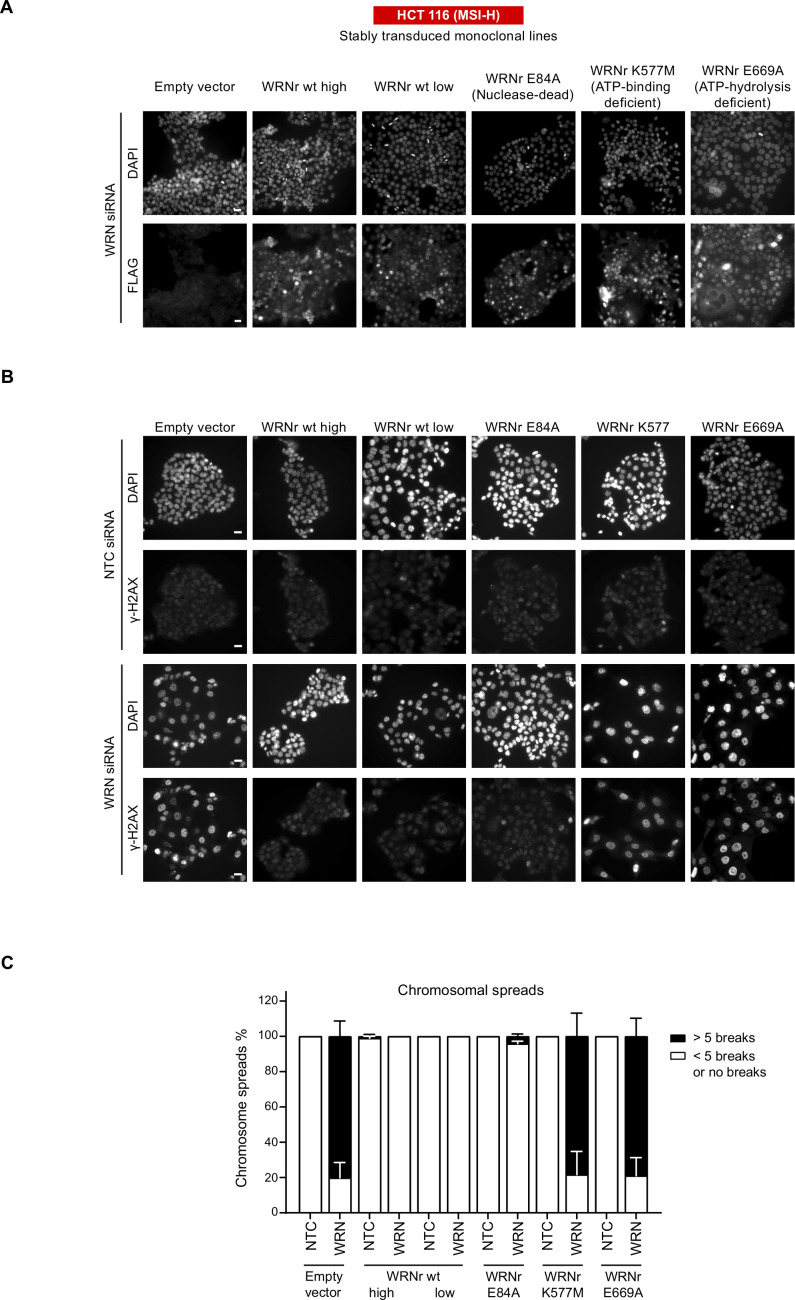
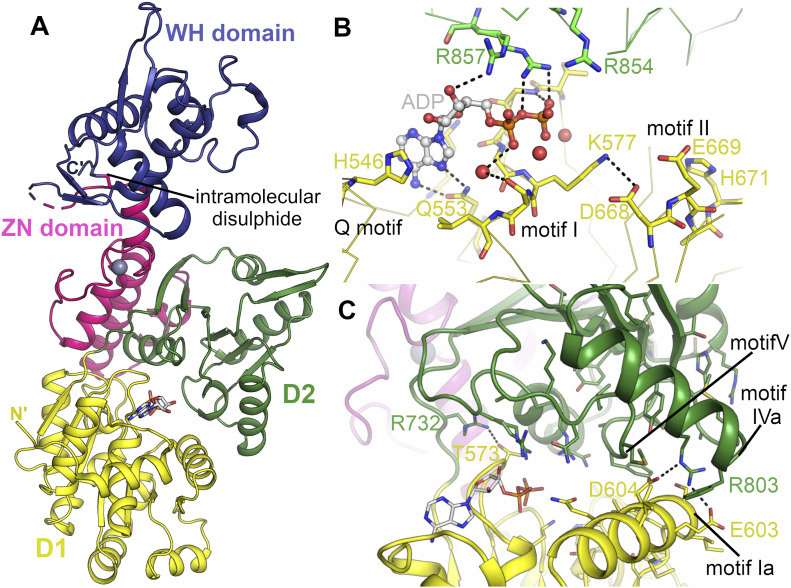
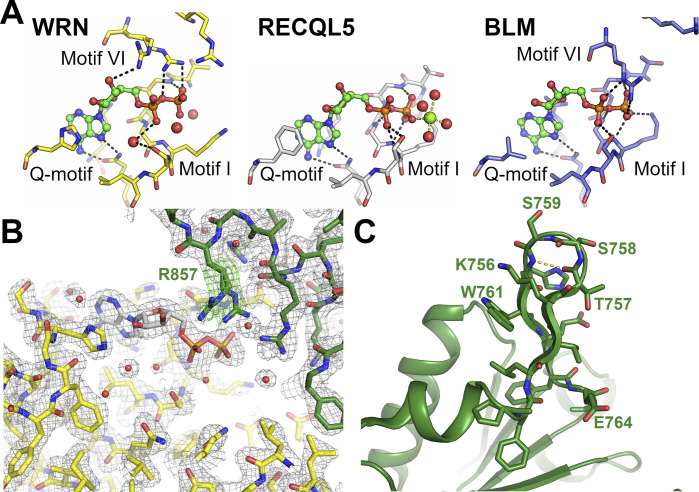
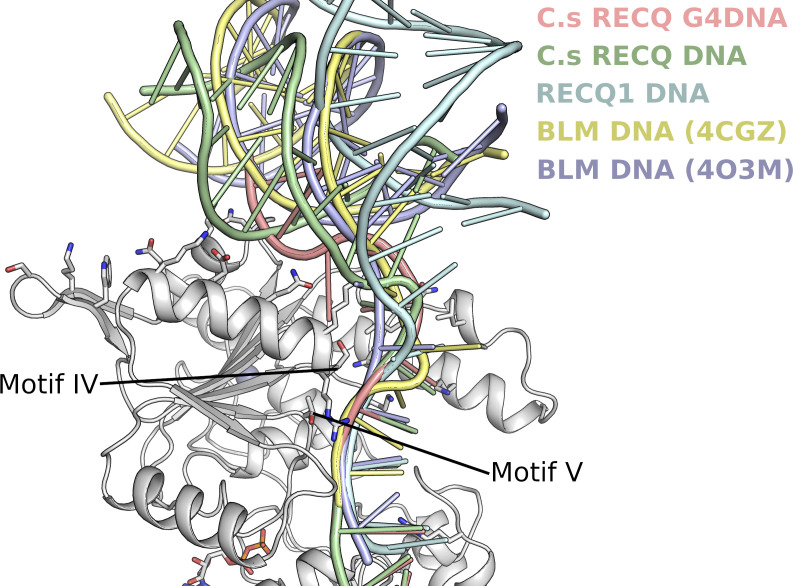
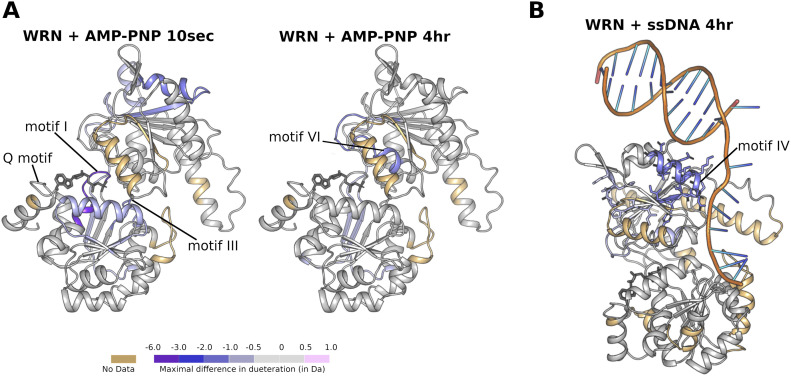
Loss of WRN, a DNA repair helicase, was identified as a strong vulnerability of microsatellite instable (MSI) cancers, making WRN a promising drug target. We show that ATP binding and hydrolysis are required for genome integrity and viability of MSI cancer cells. We report a 2.2-Å crystal structure of the WRN helicase core (517-1,093), comprising the two helicase subdomains and winged helix domain but not the HRDC domain or nuclease domains. The structure highlights unusual features. First, an atypical mode of nucleotide binding that results in unusual relative positioning of the two helicase subdomains. Second, an additional β-hairpin in the second helicase subdomain and an unusual helical hairpin in the Zn2+ binding domain. Modelling of the WRN helicase in complex with DNA suggests roles for these features in the binding of alternative DNA structures. NMR analysis shows a weak interaction between the HRDC domain and the helicase core, indicating a possible biological role for this association. Together, this study will facilitate the structure-based development of inhibitors against WRN helicase.
© 2020 Newman et al.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
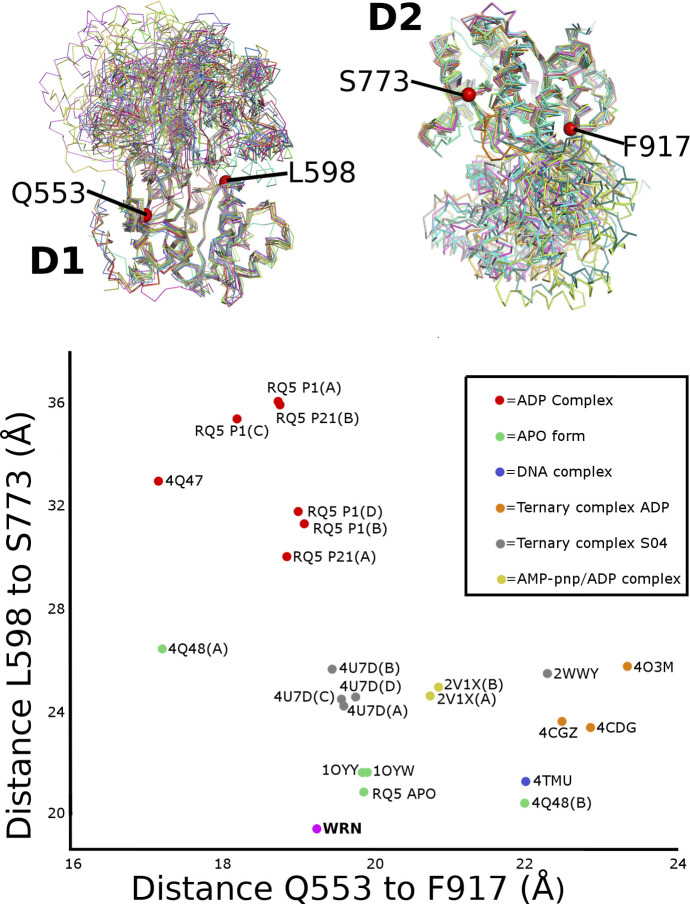
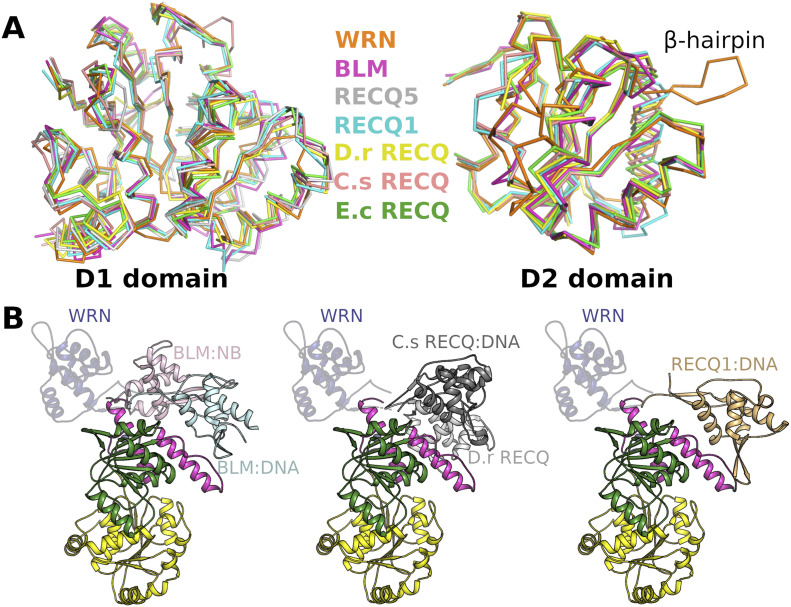
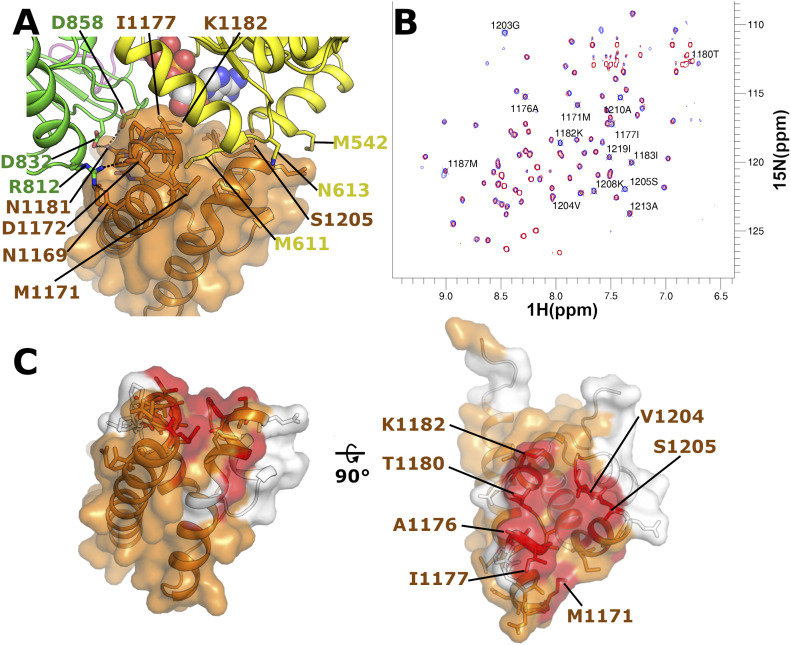
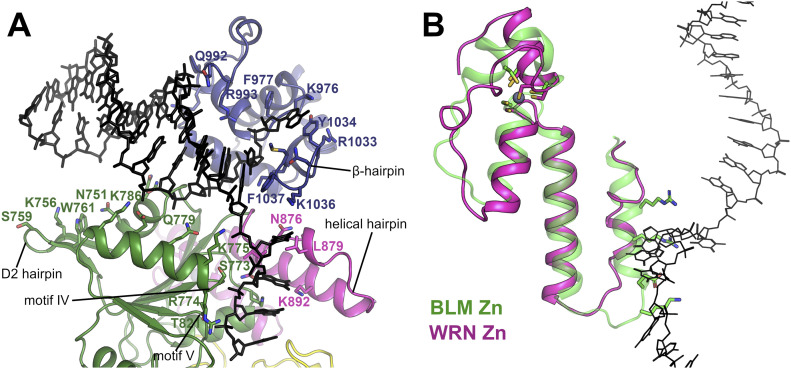
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases