

Dynamic regulation of coral energy metabolism throughout the diel cycle
- PMID: 33199772
- PMCID: PMC7669893
- DOI: 10.1038/s41598-020-76828-2
Dynamic regulation of coral energy metabolism throughout the diel cycle
Abstract
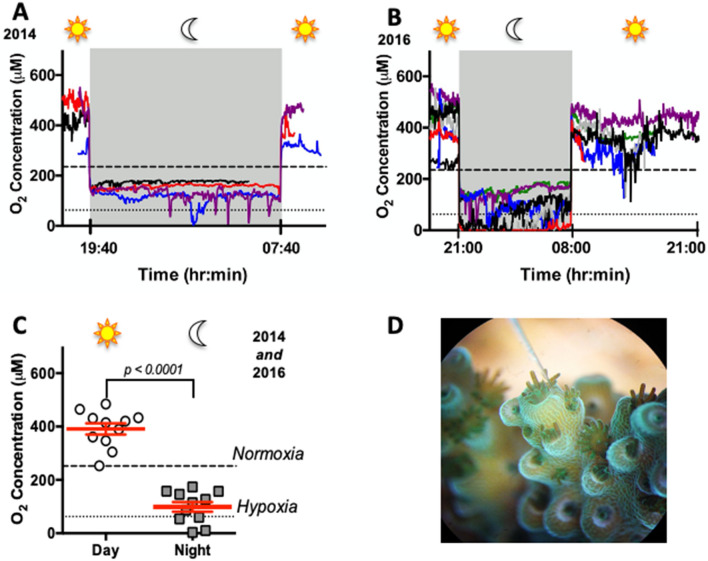
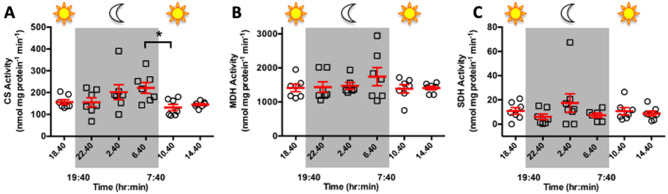
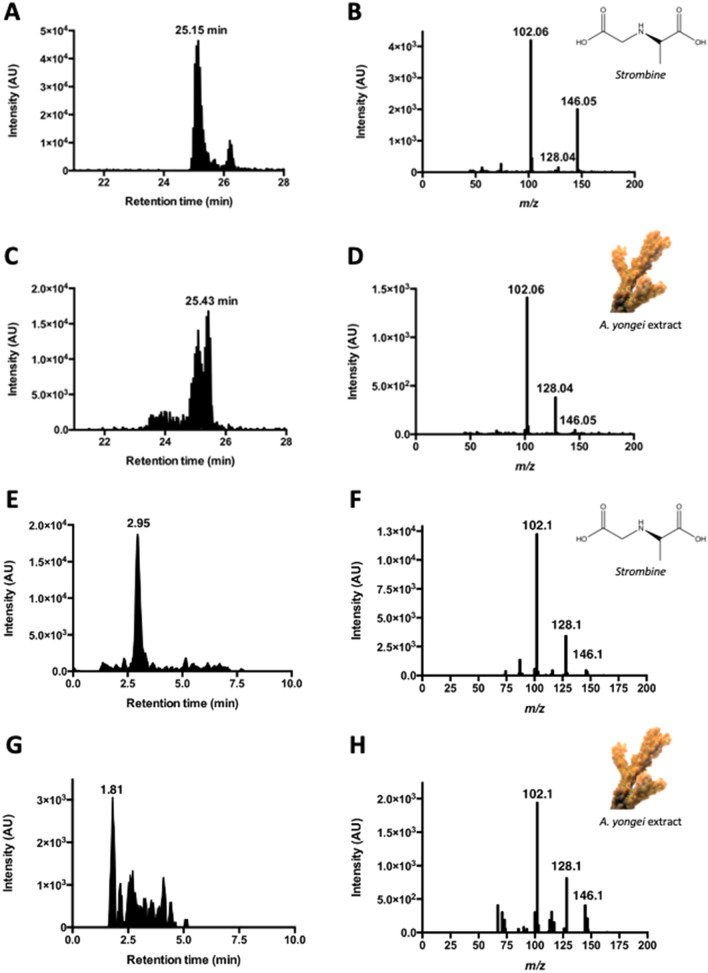
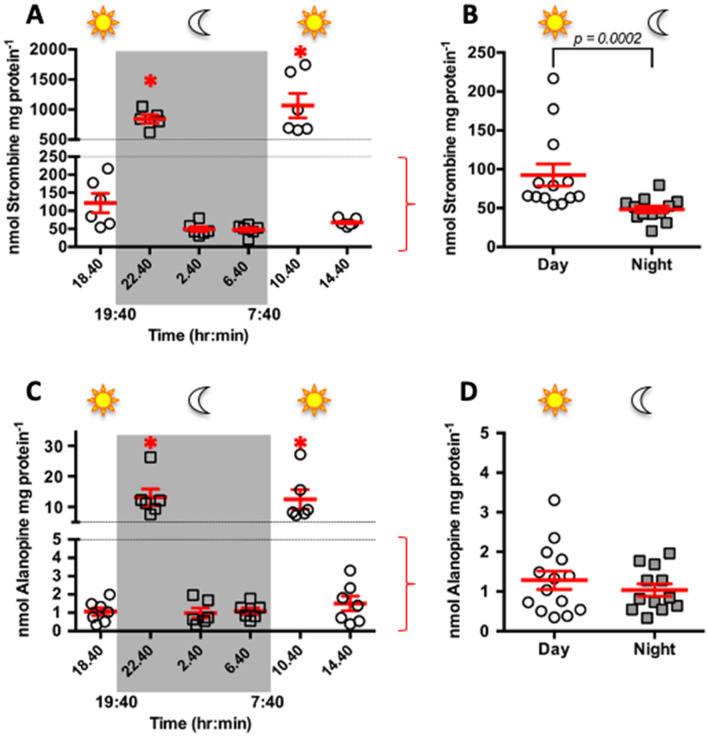
Coral reefs are naturally exposed to daily and seasonal variations in environmental oxygen levels, which can be exacerbated in intensity and duration by anthropogenic activities. However, coral's diel oxygen dynamics and fermentative pathways remain poorly understood. Here, continuous oxygen microelectrode recordings in the coral diffusive boundary layer revealed hyperoxia during daytime and hypoxia at nighttime resulting from net photosynthesis and net respiration, respectively. The activities of the metabolic enzymes citrate synthase (CS), malate dehydrogenase, and strombine dehydrogenase remained constant throughout the day/night cycle, suggesting that energy metabolism was regulated through adjustments in metabolite fluxes and not through changes in enzyme abundance. Liquid chromatography-mass spectrometry analyses identified strombine as coral's main fermentative end product. Strombine levels peaked as oxygen became depleted at dusk, indicating increased fermentation rates at the onset of nightly hypoxia, and again at dawn as photosynthesis restored oxygen and photosynthate supply. When these peaks were excluded from the analyses, average strombine levels during the day were nearly double those at night, indicating sifnificant fermentation rates even during aerobic conditions. These results highlight the dynamic changes in oxygen levels in the coral diffusive boundary layer, and the importance of fermentative metabolism for coral biology.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Effects of bleaching on oxygen dynamics and energy metabolism of two Caribbean coral species.Sci Total Environ. 2024 Apr 1;919:170753. doi: 10.1016/j.scitotenv.2024.170753. Epub 2024 Feb 13. Sci Total Environ. 2024. PMID: 38360316
-
Mutualistic damselfish induce higher photosynthetic rates in their host coral.J Exp Biol. 2017 May 15;220(Pt 10):1803-1811. doi: 10.1242/jeb.152462. J Exp Biol. 2017. PMID: 28515171
-
In situ coral reef oxygen metabolism: an eddy correlation study.PLoS One. 2013;8(3):e58581. doi: 10.1371/journal.pone.0058581. Epub 2013 Mar 11. PLoS One. 2013. PMID: 23536798 Free PMC article.
-
Coral photobiology: new light on old views.Zoology (Jena). 2015 Apr;118(2):71-8. doi: 10.1016/j.zool.2014.08.003. Epub 2014 Nov 3. Zoology (Jena). 2015. PMID: 25467066 Review.
-
Warm-water coral reefs and climate change.Science. 2015 Nov 13;350(6262):769-71. doi: 10.1126/science.aad0349. Science. 2015. PMID: 26564846 Review.
Cited by
-
DNA from non-viable bacteria biases diversity estimates in the corals Acropora loripes and Pocillopora acuta.Environ Microbiome. 2023 Dec 8;18(1):86. doi: 10.1186/s40793-023-00541-6. Environ Microbiome. 2023. PMID: 38062479 Free PMC article.
-
Intra-colony spatial variance of oxyregulation and hypoxic thresholds for key Acropora coral species.Ecol Evol. 2024 Mar 5;14(3):e11100. doi: 10.1002/ece3.11100. eCollection 2024 Mar. Ecol Evol. 2024. PMID: 38444722 Free PMC article.
-
Spatiotemporal variability of oxygen concentration in coral reefs of Gorgona Island (Eastern Tropical Pacific) and its effect on the coral Pocillopora capitata.PeerJ. 2023 Jan 26;11:e14586. doi: 10.7717/peerj.14586. eCollection 2023. PeerJ. 2023. PMID: 36721774 Free PMC article.
-
Bacterial regulation of coral larval metamorphosis and settlement in Pocillopora damicornis.Commun Biol. 2025 Aug 23;8(1):1271. doi: 10.1038/s42003-025-08720-6. Commun Biol. 2025. PMID: 40849345 Free PMC article.
-
An Organismal Perspective on the Warburg Effect and Models for Proliferation Studies.Biology (Basel). 2023 Mar 26;12(4):502. doi: 10.3390/biology12040502. Biology (Basel). 2023. PMID: 37106703 Free PMC article. Review.
References
-
- Ohde S, van Woesik R. Carbon dioxide flux and metabolic processes of a coral reef, Okinawa. Bull. Mar. Sci. 1999;65:559–576.
-
- Smith JE, Price NN, Nelson CE, Haas AF. Coupled changes in oxygen concentration and pH caused by metabolism of benthic coral reef organisms. Mar. Biol. 2013;160:2437–2447. doi: 10.1007/s00227-013-2239-z. - DOI
-
- Kühl M, Cohen Y, Dalsgaard T, Jorgensen BB, Revbech NP. Microenvironment and photosynthesis of zooxanthellae in scleractinian corals studied with microsensors for O2, pH and light. Mar. Ecol. Prog. Ser. 1995;117:159–172. doi: 10.3354/meps117159. - DOI
