

Structural and functional consequences of reversible lipid asymmetry in living membranes
- PMID: 33199908
- PMCID: PMC7747298
- DOI: 10.1038/s41589-020-00688-0
Structural and functional consequences of reversible lipid asymmetry in living membranes
Abstract
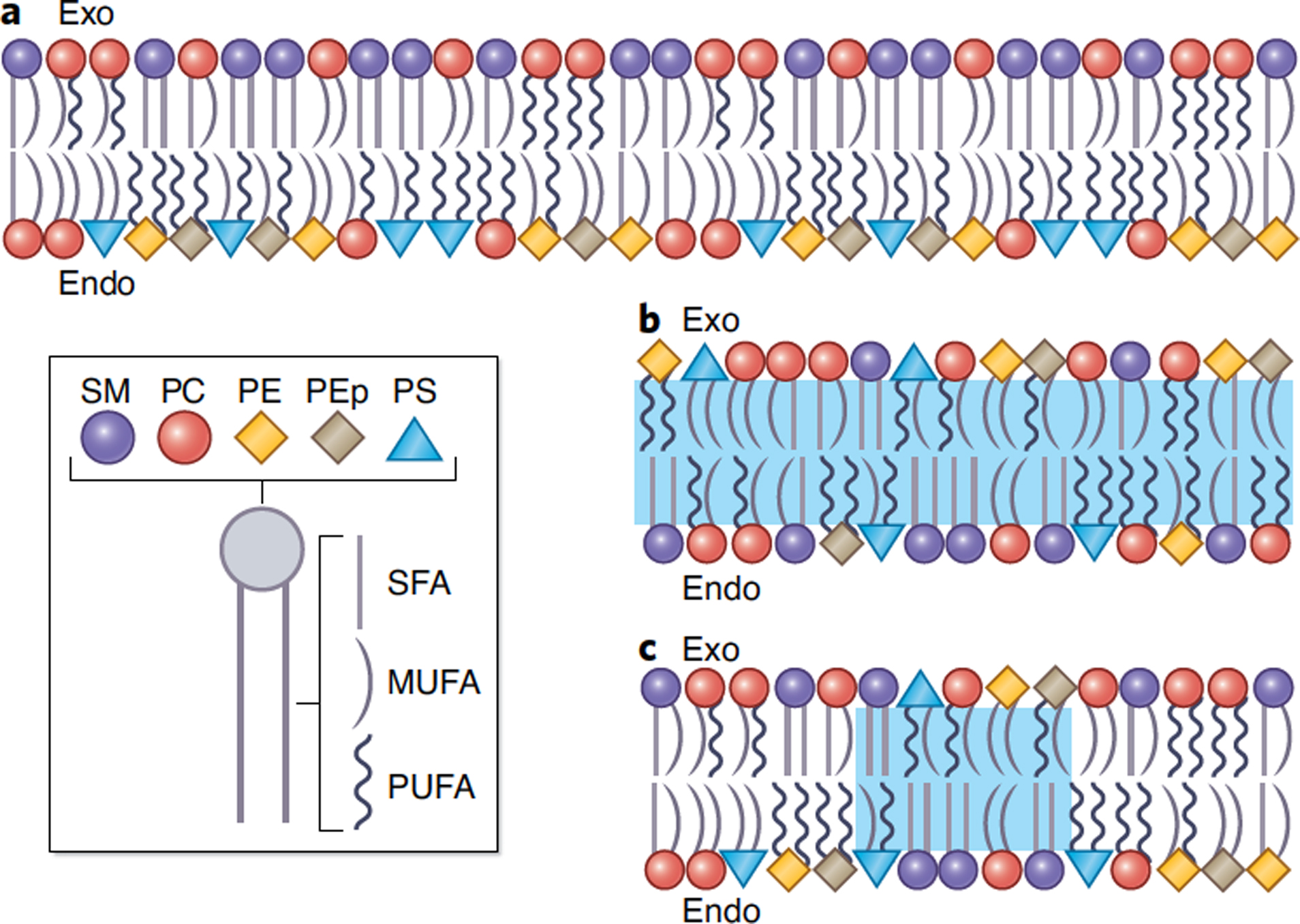
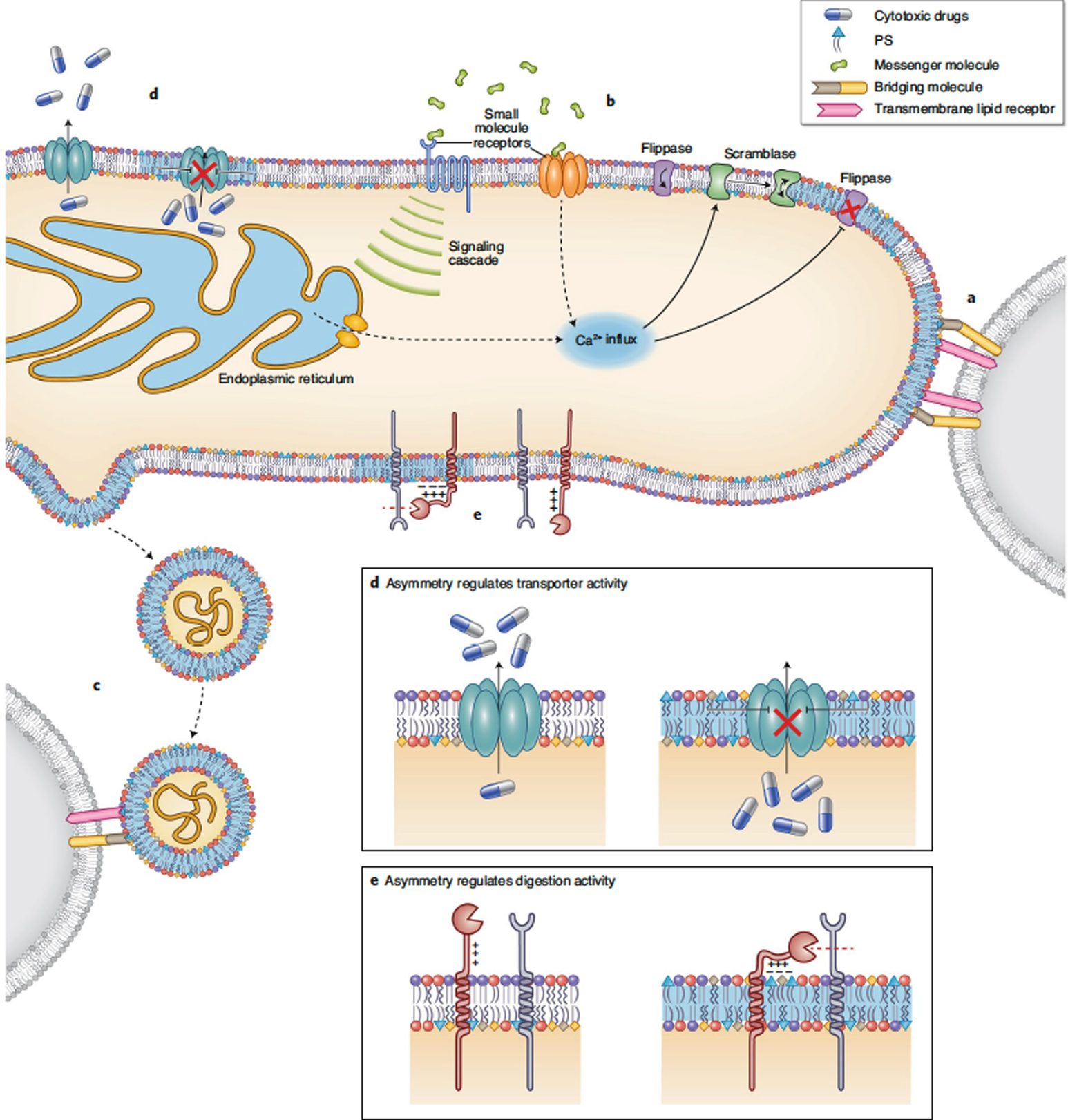
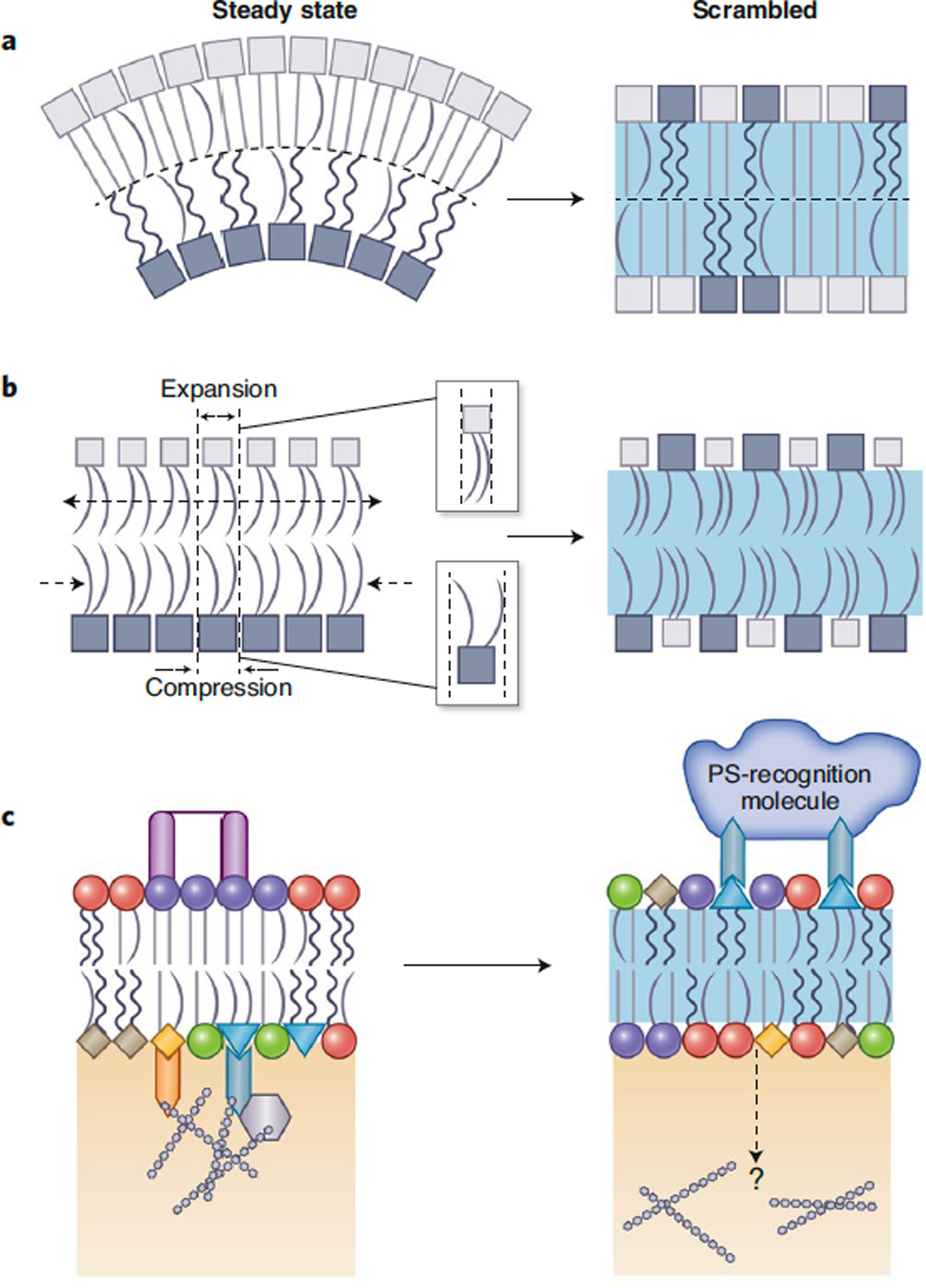
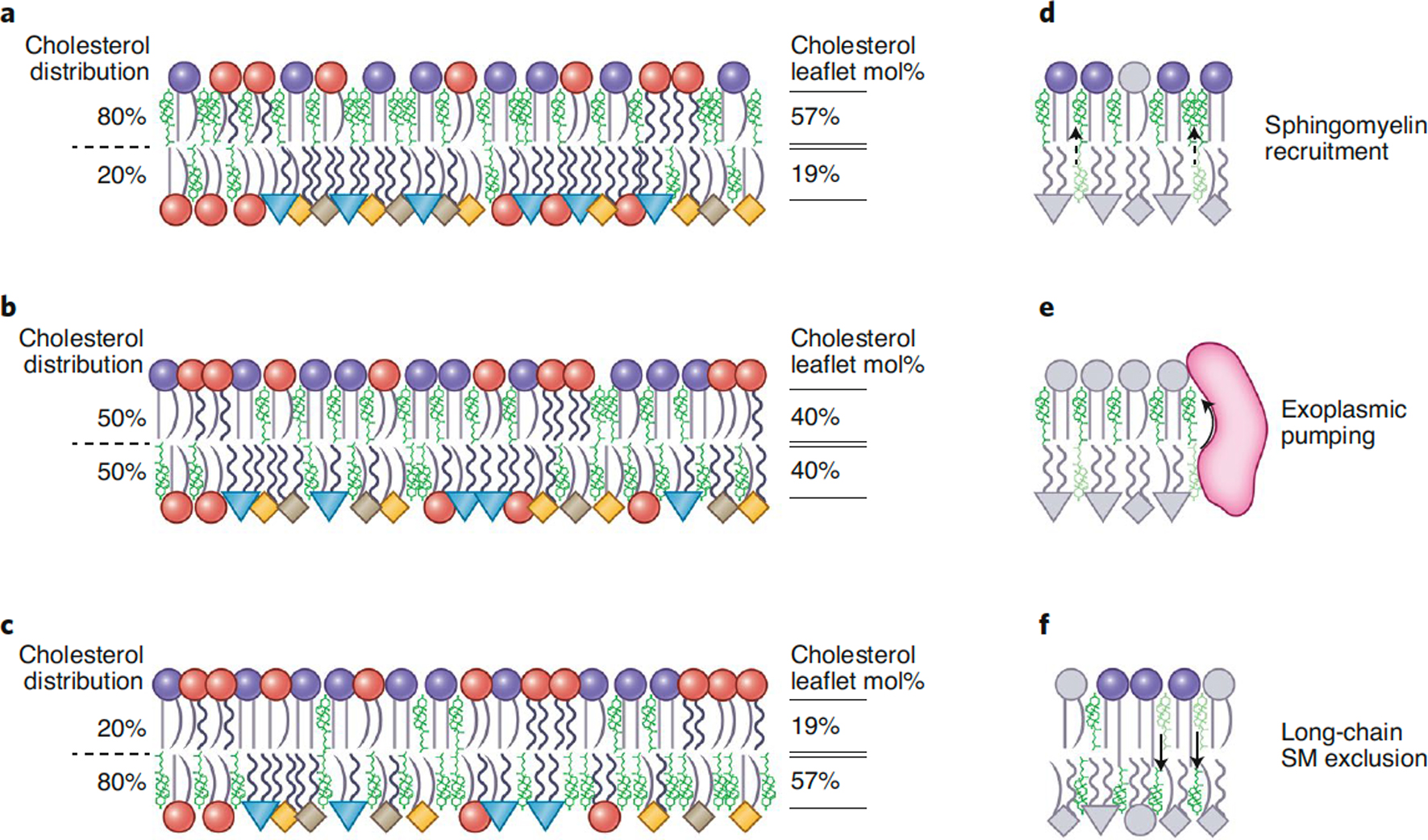
Maintenance of lipid asymmetry across the two leaflets of the plasma membrane (PM) bilayer is a ubiquitous feature of eukaryotic cells. Loss of this asymmetry has been widely associated with cell death. However, increasing evidence points to the physiological importance of non-apoptotic, transient changes in PM asymmetry. Such transient scrambling events are associated with a range of biological functions, including intercellular communication and intracellular signaling. Thus, regulation of interleaflet lipid distribution in the PM is a broadly important but underappreciated cellular process with key physiological and structural consequences. Here, we compile the mounting evidence revealing multifaceted, functional roles of PM asymmetry and transient loss thereof. We discuss the consequences of reversible asymmetry on PM structure, biophysical properties and interleaflet coupling. We argue that despite widespread recognition of broad aspects of membrane asymmetry, its importance in cell biology demands more in-depth investigation of its features, regulation, and physiological and pathological implications.
Figures
Similar articles
-
Understanding the Link between Lipid Diversity and the Biophysical Properties of the Neuronal Plasma Membrane.Biochemistry. 2020 Aug 25;59(33):3010-3018. doi: 10.1021/acs.biochem.0c00524. Epub 2020 Aug 12. Biochemistry. 2020. PMID: 32786397
-
Multivalent lipid targeting by the calcium-independent C2A domain of synaptotagmin-like protein 4/granuphilin.J Biol Chem. 2021 Jan-Jun;296:100159. doi: 10.1074/jbc.RA120.014618. Epub 2020 Dec 10. J Biol Chem. 2021. PMID: 33277360 Free PMC article.
-
Influence of membrane phospholipid composition and structural organization on spontaneous lipid transfer between membranes.Gen Physiol Biophys. 2006 Sep;25(3):313-24. Gen Physiol Biophys. 2006. PMID: 17197729
-
Phospholipid unsaturation and plasma membrane organization.Chem Phys Lipids. 1975 May;14(3):236-46. doi: 10.1016/0009-3084(75)90005-5. Chem Phys Lipids. 1975. PMID: 165015 Review.
-
The Lipids of the Early Endosomes: Making Multimodality Work.Chembiochem. 2017 Jun 19;18(12):1053-1060. doi: 10.1002/cbic.201700046. Epub 2017 May 10. Chembiochem. 2017. PMID: 28374483 Review.
Cited by
-
Eugene P. Kennedy's Legacy: Defining Bacterial Phospholipid Pathways and Function.Front Mol Biosci. 2021 Mar 25;8:666203. doi: 10.3389/fmolb.2021.666203. eCollection 2021. Front Mol Biosci. 2021. PMID: 33842554 Free PMC article. Review.
-
Age and sex are associated with the plasma lipidome: findings from the GOLDN study.Lipids Health Dis. 2021 Apr 3;20(1):30. doi: 10.1186/s12944-021-01456-2. Lipids Health Dis. 2021. PMID: 33812378 Free PMC article. Clinical Trial.
-
Native-like environments afford novel mechanistic insights into membrane proteins.Trends Biochem Sci. 2022 Jul;47(7):561-569. doi: 10.1016/j.tibs.2022.02.008. Epub 2022 Mar 21. Trends Biochem Sci. 2022. PMID: 35331611 Free PMC article. Review.
-
Nonconverged Constraints Cause Artificial Temperature Gradients in Lipid Bilayer Simulations.J Phys Chem B. 2021 Aug 26;125(33):9537-9546. doi: 10.1021/acs.jpcb.1c03665. Epub 2021 Aug 16. J Phys Chem B. 2021. PMID: 34398598 Free PMC article.
-
Distributing aminophospholipids asymmetrically across leaflets causes anomalous membrane stiffening.Biophys J. 2023 Jun 20;122(12):2445-2455. doi: 10.1016/j.bpj.2023.04.025. Epub 2023 Apr 29. Biophys J. 2023. PMID: 37120716 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
