

Precise control of ion channel and gap junction expression is required for patterning of the regenerating axolotl limb
- PMID: 33200809
- PMCID: PMC8796139
- DOI: 10.1387/ijdb.200114jw
Precise control of ion channel and gap junction expression is required for patterning of the regenerating axolotl limb
Abstract
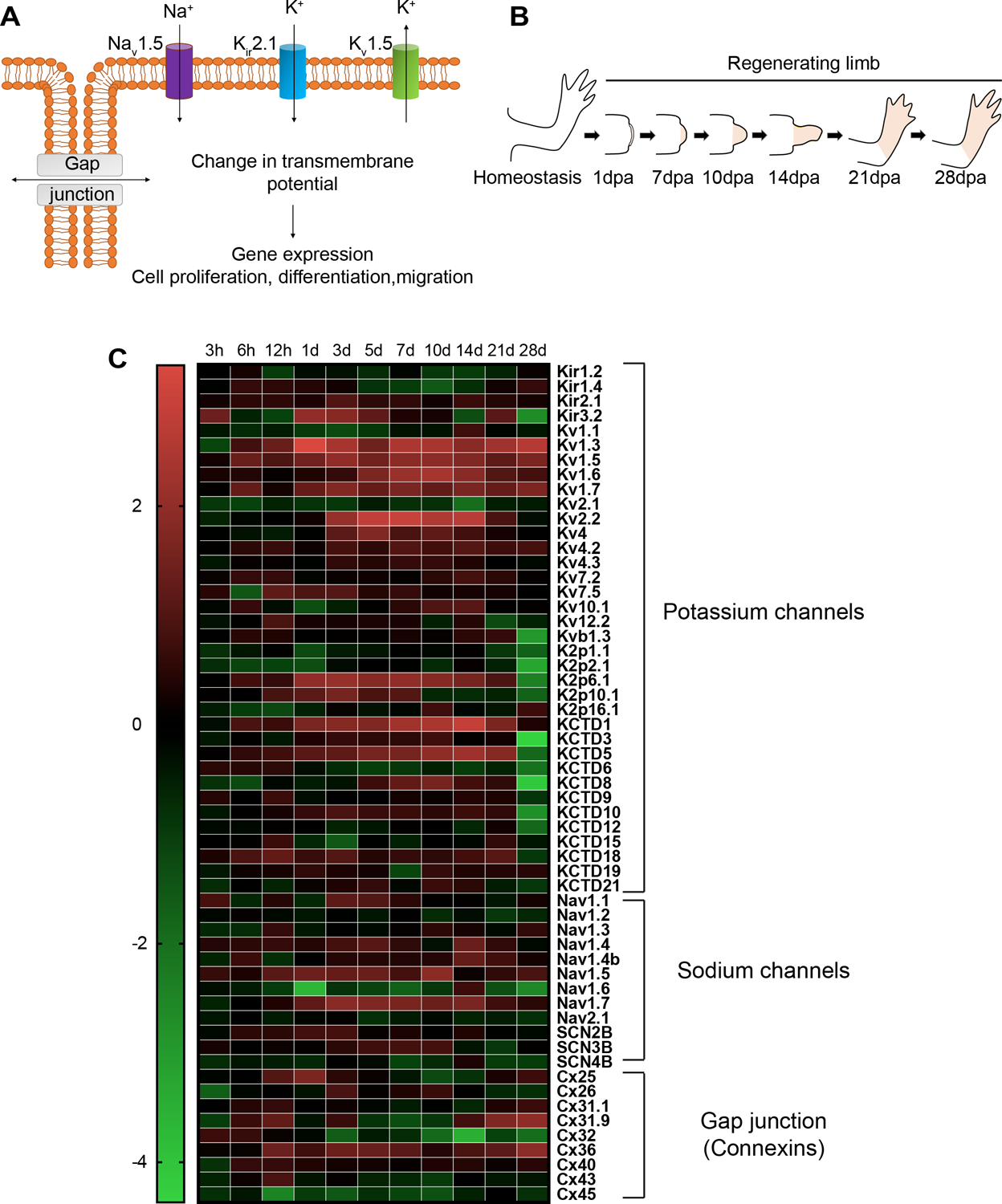
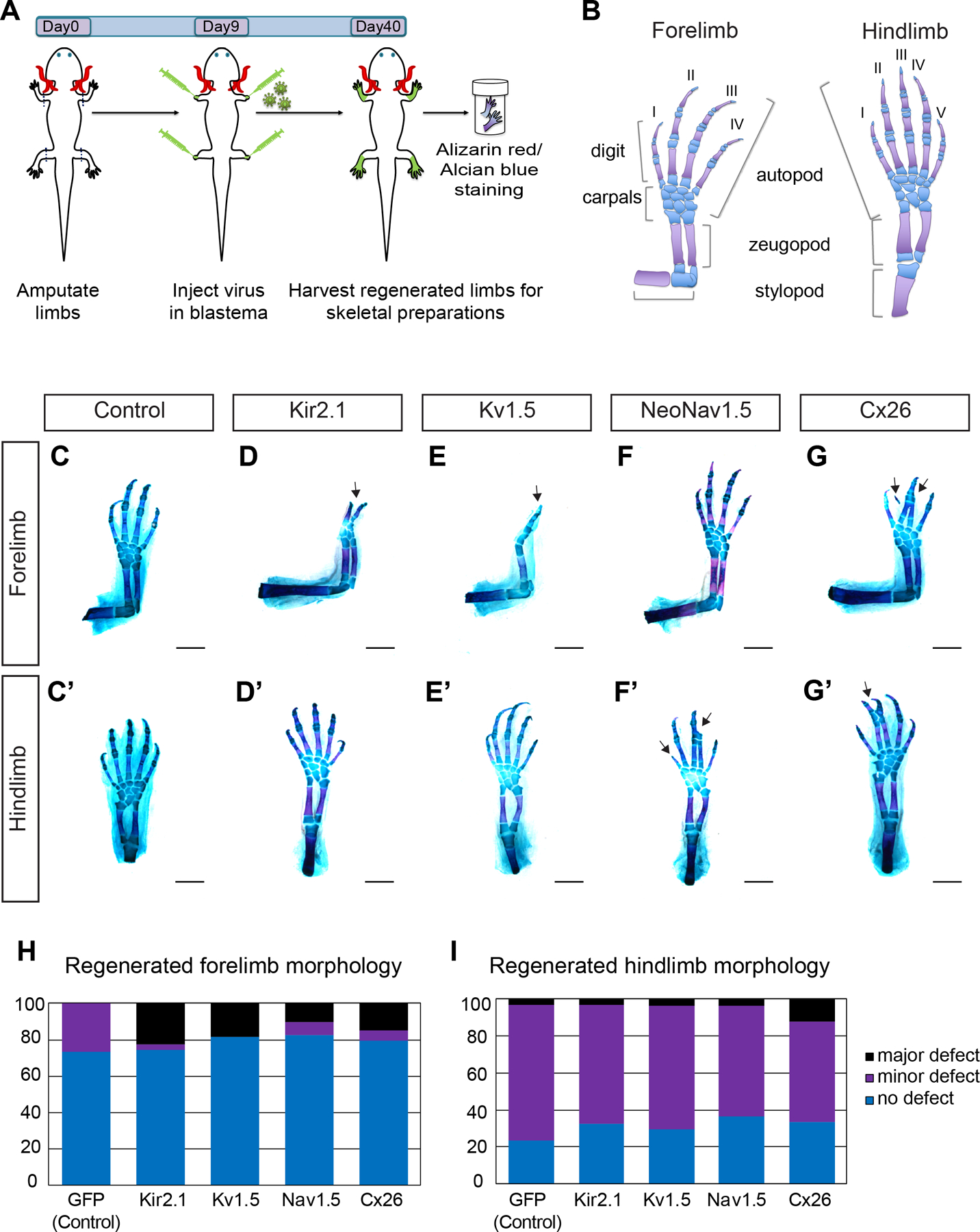
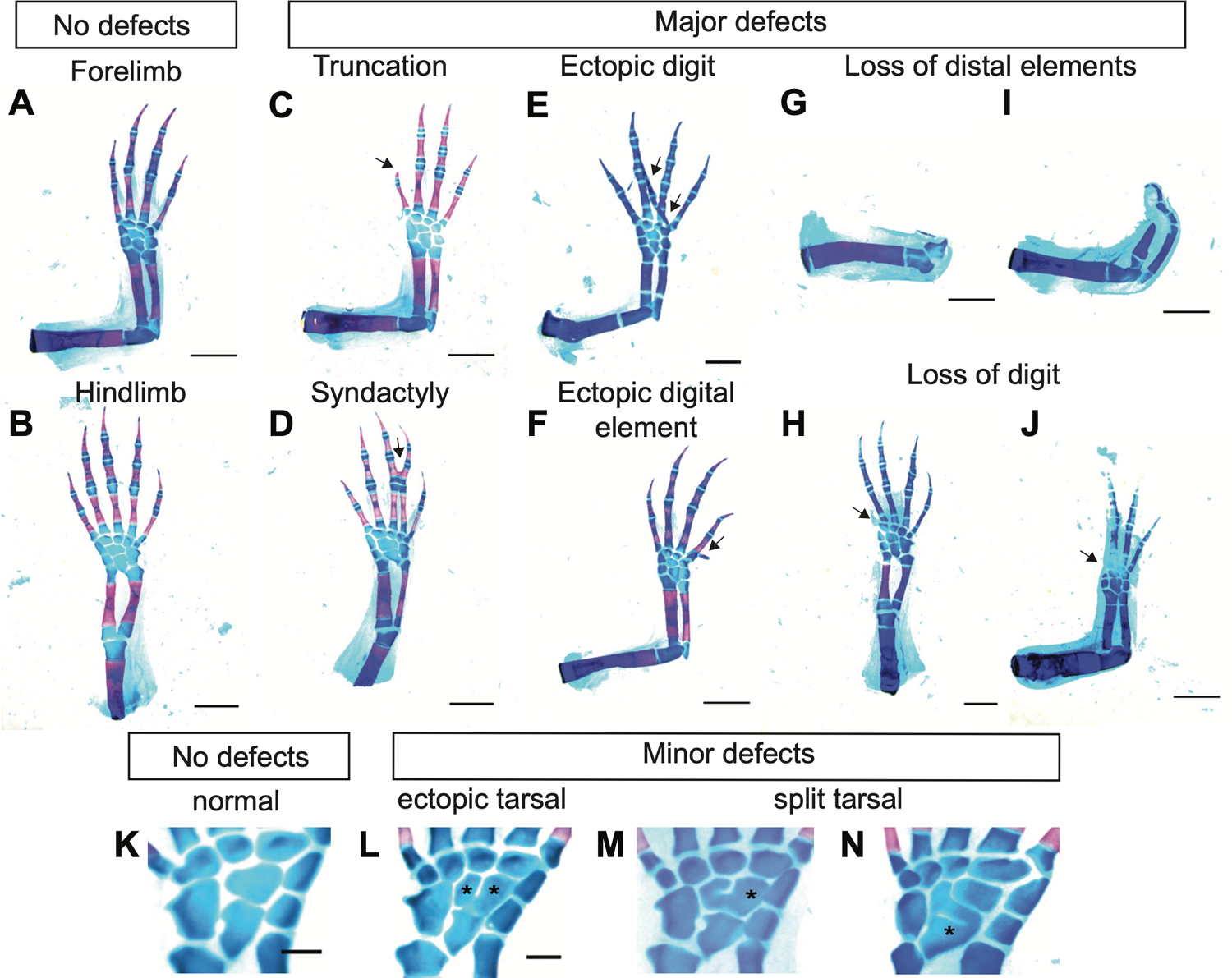
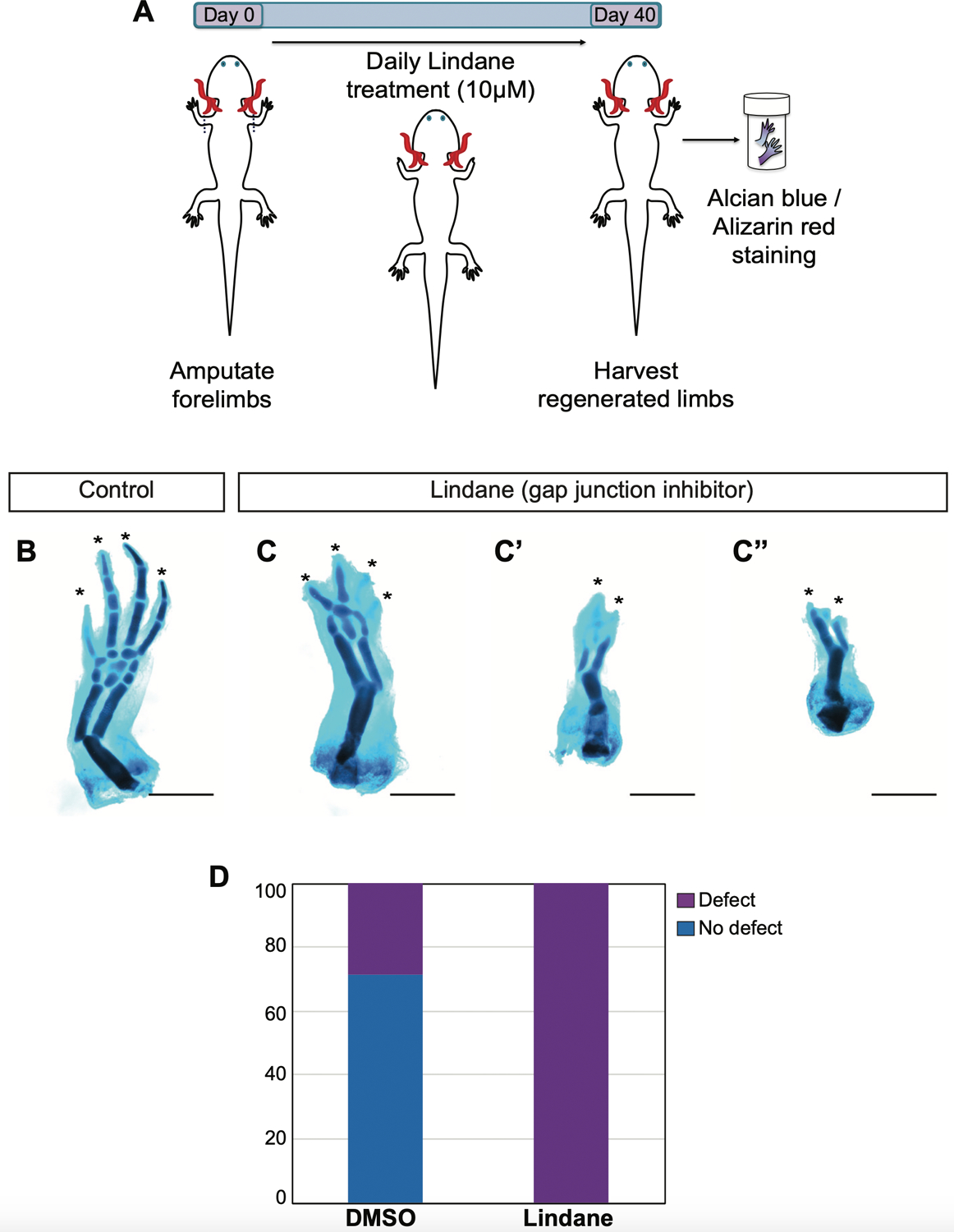
Axolotls and other salamanders have the capacity to regenerate lost tissue after an amputation or injury. Growth and morphogenesis are coordinated within cell groups in many contexts by the interplay of transcriptional networks and biophysical properties such as ion flows and voltage gradients. It is not, however, known whether regulators of a cell's ionic state are involved in limb patterning at later stages of regeneration. Here we manipulated expression and activities of ion channels and gap junctions in vivo, in axolotl limb blastema cells. Limb amputations followed by retroviral infections were performed to drive expression of a human gap junction protein Connexin 26 (Cx26), potassium (Kir2.1-Y242F and Kv1.5) and sodium (NeoNav1.5) ion channel proteins along with EGFP control. Skeletal preparation revealed that overexpressing Cx26 caused syndactyly, while overexpression of ion channel proteins resulted in digit loss and structural abnormalities compared to EGFP expressing control limbs. Additionally, we showed that exposing limbs to the gap junction inhibitor lindane during the regeneration process caused digit loss. Our data reveal that manipulating native ion channel and gap junction function in blastema cells results in patterning defects involving the number and structure of the regenerated digits. Gap junctions and ion channels have been shown to mediate ion flows that control the endogenous voltage gradients which are tightly associated with the regulation of gene expression, cell cycle progression, migration, and other cellular behaviors. Therefore, we postulate that mis-expression of these channels may have disturbed this regulation causing uncoordinated cell behavior which results in morphological defects.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- ADAMS DS, MASI A & LEVIN M (2007). H+ pump-dependent changes in membrane voltage are an early mechanism necessary and sufficient to induce Xenopus tail regeneration. Development, 134: 1323–35. - PubMed
-
- BATES E (2015). Ion Channels in Development and Cancer. Annu Rev Cell Dev Biol, 31: 231–47. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
