

G ATA2 mediates the negative regulation of the prepro-thyrotropin-releasing hormone gene by liganded T3 receptor β2 in the rat hypothalamic paraventricular nucleus
- PMID: 33201916
- PMCID: PMC7671546
- DOI: 10.1371/journal.pone.0242380
G ATA2 mediates the negative regulation of the prepro-thyrotropin-releasing hormone gene by liganded T3 receptor β2 in the rat hypothalamic paraventricular nucleus
Abstract
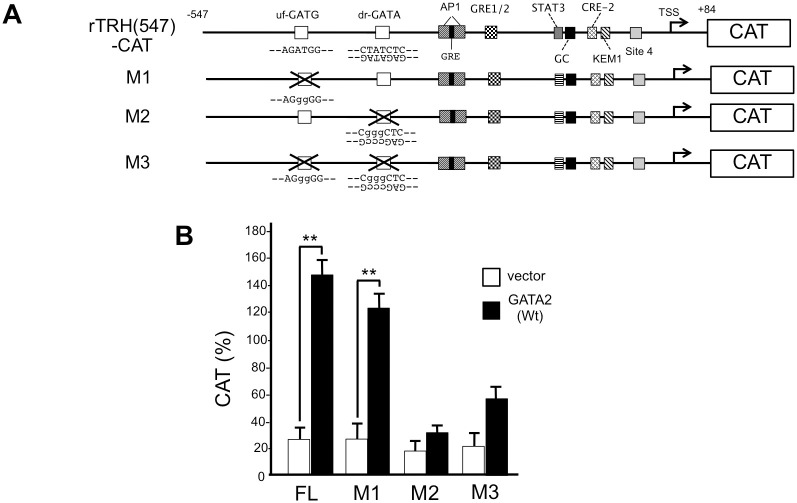
Thyroid hormone (T3) inhibits thyrotropin-releasing hormone (TRH) synthesis in the hypothalamic paraventricular nucleus (PVN). Although the T3 receptor (TR) β2 is known to mediate the negative regulation of the prepro-TRH gene, its molecular mechanism remains unknown. Our previous studies on the T3-dependent negative regulation of the thyrotropin β subunit (TSHβ) gene suggest that there is a tethering mechanism, whereby liganded TRβ2 interferes with the function of the transcription factor, GATA2, a critical activator of the TSHβ gene. Interestingly, the transcription factors Sim1 and Arnt2, the determinants of PVN differentiation in the hypothalamus, are reported to induce expression of TRβ2 and GATA2 in cultured neuronal cells. Here, we confirmed the expression of the GATA2 protein in the TRH neuron of the rat PVN using immunohistochemistry with an anti-GATA2 antibody. According to an experimental study from transgenic mice, a region of the rat prepro-TRH promoter from nt. -547 to nt. +84 was able to mediate its expression in the PVN. We constructed a chloramphenicol acetyltransferase (CAT) reporter gene containing this promoter sequence (rTRH(547)-CAT) and showed that GATA2 activated the promoter in monkey kidney-derived CV1 cells. Deletion and mutation analyses identified a functional GATA-responsive element (GATA-RE) between nt. -357 and nt. -352. When TRβ2 was co-expressed, T3 reduced GATA2-dependent promoter activity to approximately 30%. Unexpectedly, T3-dependent negative regulation was maintained after mutation of the reported negative T3-responsive element, site 4. T3 also inhibited the GATA2-dependent transcription enhanced by cAMP agonist, 8-bromo-cAMP. A rat thyroid medullary carcinoma cell line, CA77, is known to express the preproTRH mRNA. Using a chromatin immunoprecipitation assay with this cell line where GATA2 expression plasmid was transfected, we observed the recognition of the GATA-RE by GATA2. We also confirmed GATA2 binding using gel shift assay with the probe for the GATA-RE. In CA77 cells, the activity of rTRH(547)-CAT was potentiated by overexpression of GATA2, and it was inhibited in a T3-dependent manner. These results suggest that GATA2 transactivates the rat prepro-TRH gene and that liganded TRβ2 interferes with this activation via a tethering mechanism as in the case of the TSHβ gene.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures







Similar articles
-
Liganded T3 receptor β2 inhibits the positive feedback autoregulation of the gene for GATA2, a transcription factor critical for thyrotropin production.PLoS One. 2020 Jan 15;15(1):e0227646. doi: 10.1371/journal.pone.0227646. eCollection 2020. PLoS One. 2020. PMID: 31940421 Free PMC article.
-
Essential Role of GATA2 in the Negative Regulation of Type 2 Deiodinase Gene by Liganded Thyroid Hormone Receptor β2 in Thyrotroph.PLoS One. 2015 Nov 16;10(11):e0142400. doi: 10.1371/journal.pone.0142400. eCollection 2015. PLoS One. 2015. PMID: 26571013 Free PMC article.
-
GATA2 mediates thyrotropin-releasing hormone-induced transcriptional activation of the thyrotropin β gene.PLoS One. 2011 Apr 14;6(4):e18667. doi: 10.1371/journal.pone.0018667. PLoS One. 2011. PMID: 21533184 Free PMC article.
-
The Mechanism of Negative Transcriptional Regulation by Thyroid Hormone: Lessons From the Thyrotropin β Subunit Gene.Vitam Horm. 2018;106:97-127. doi: 10.1016/bs.vh.2017.06.006. Epub 2017 Aug 18. Vitam Horm. 2018. PMID: 29407449 Review.
-
Feedback regulation of thyrotropin-releasing hormone gene expression by thyroid hormone in the hypothalamic paraventricular nucleus.Ciba Found Symp. 1992;168:144-58; discussion 158-64. doi: 10.1002/9780470514283.ch10. Ciba Found Symp. 1992. PMID: 1425022 Review.
Cited by
-
Impairment of the Hypothalamus-Pituitary-Thyroid Axis Caused by Naturally Occurring GATA2 Mutations In Vitro.Int J Mol Sci. 2021 Sep 16;22(18):10015. doi: 10.3390/ijms221810015. Int J Mol Sci. 2021. PMID: 34576178 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
