

Gene- and Species-Specific Hox mRNA Translation by Ribosome Expansion Segments
- PMID: 33202249
- PMCID: PMC7769145
- DOI: 10.1016/j.molcel.2020.10.023
Gene- and Species-Specific Hox mRNA Translation by Ribosome Expansion Segments
Abstract
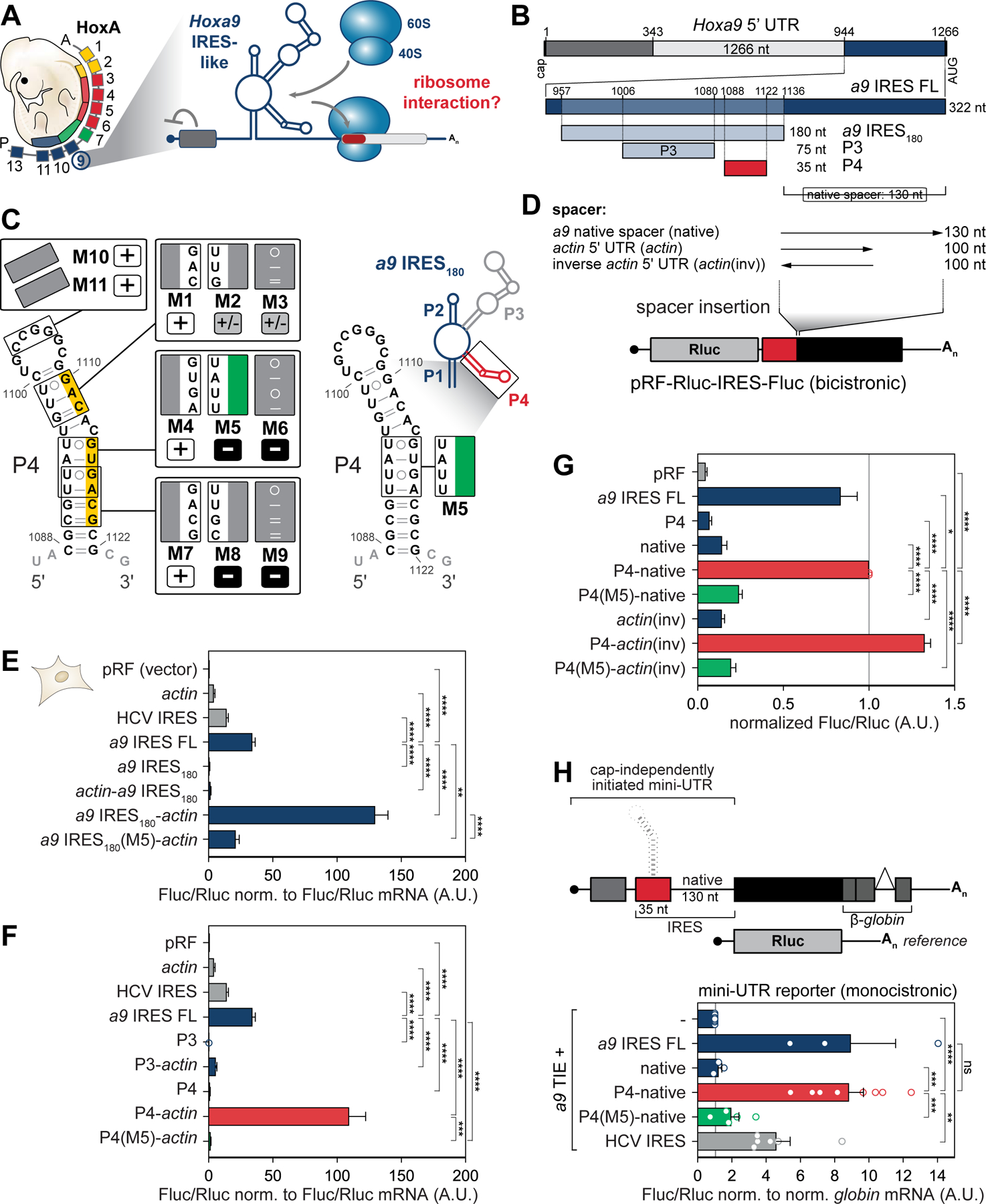
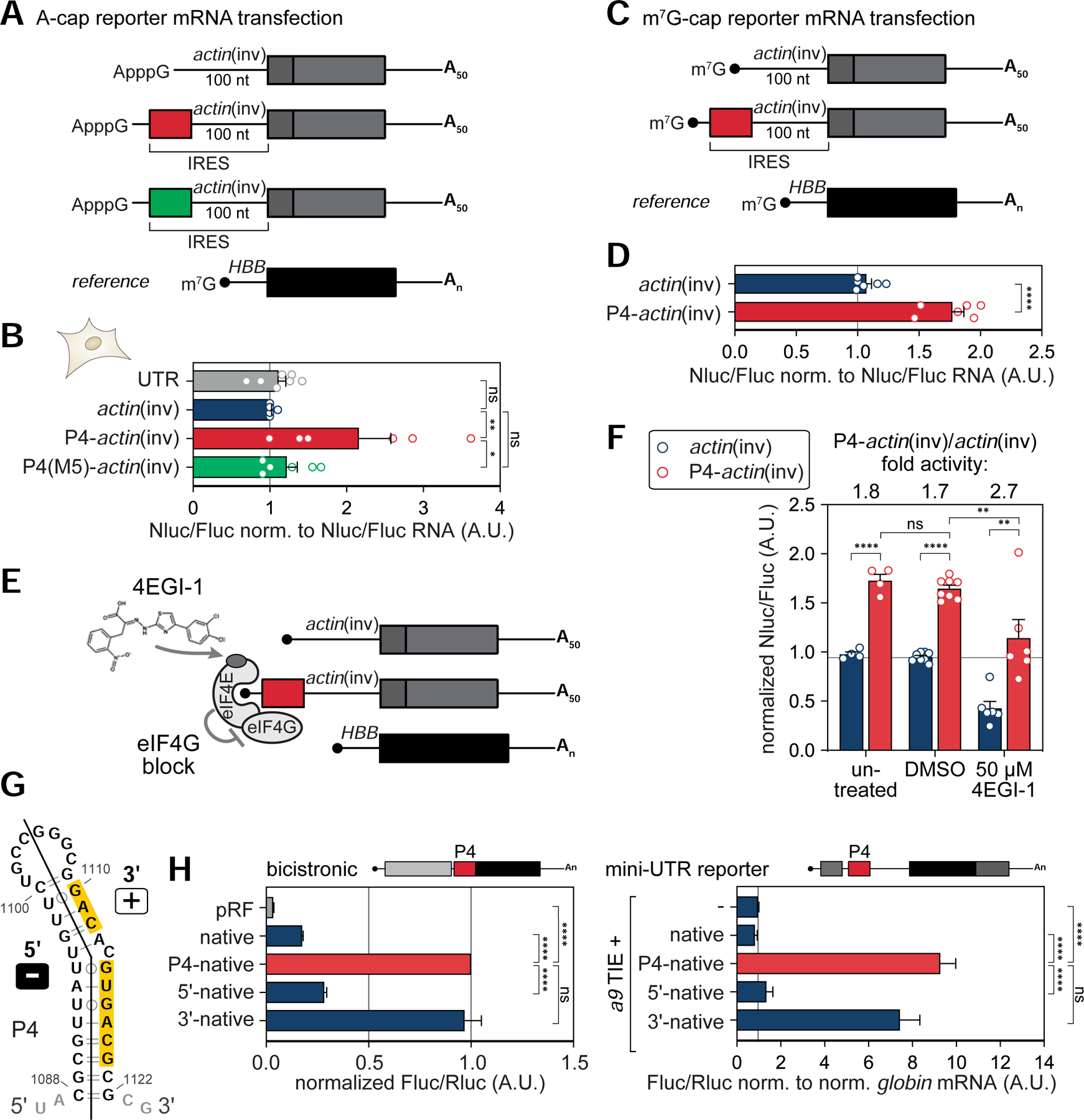
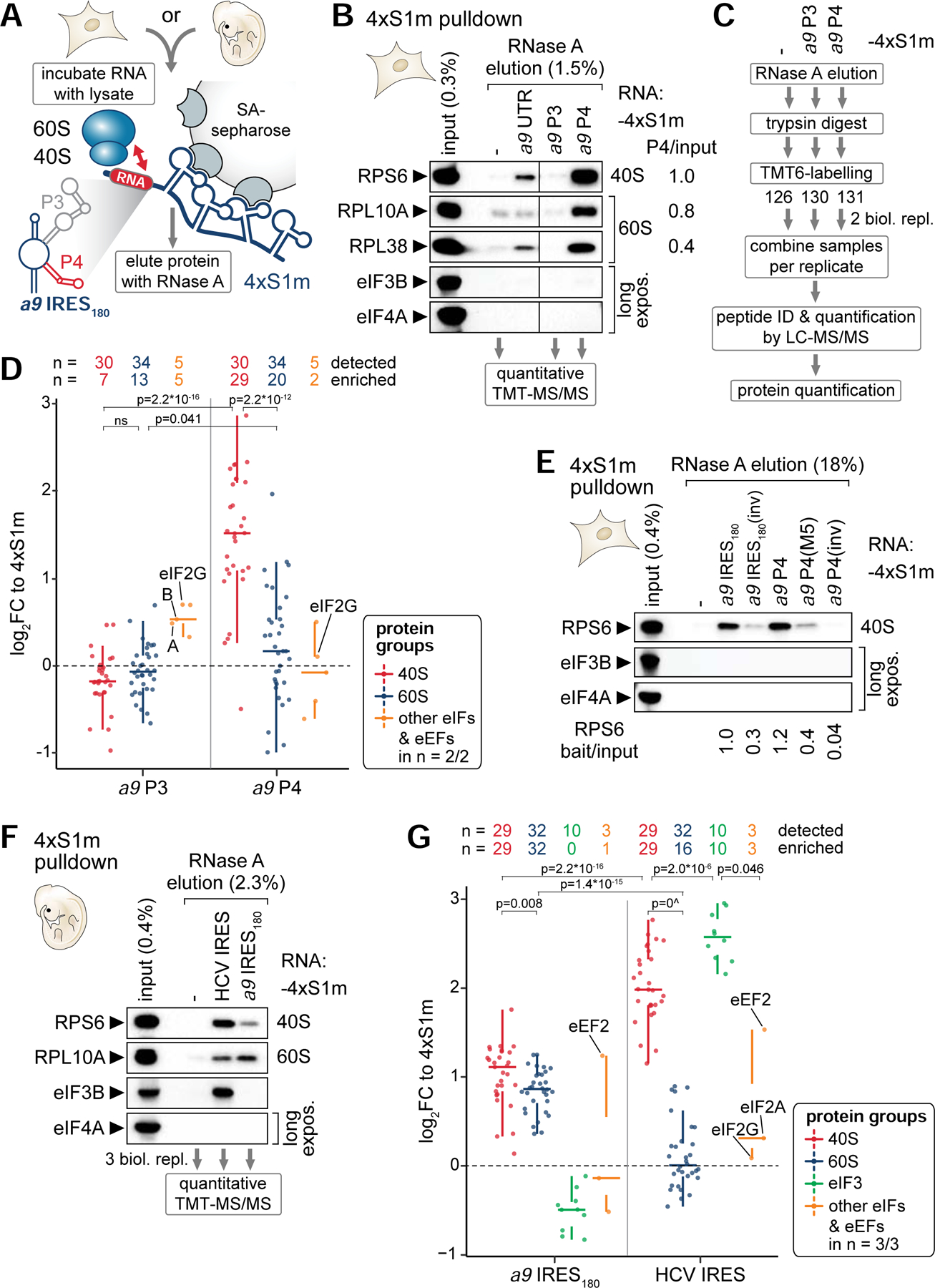
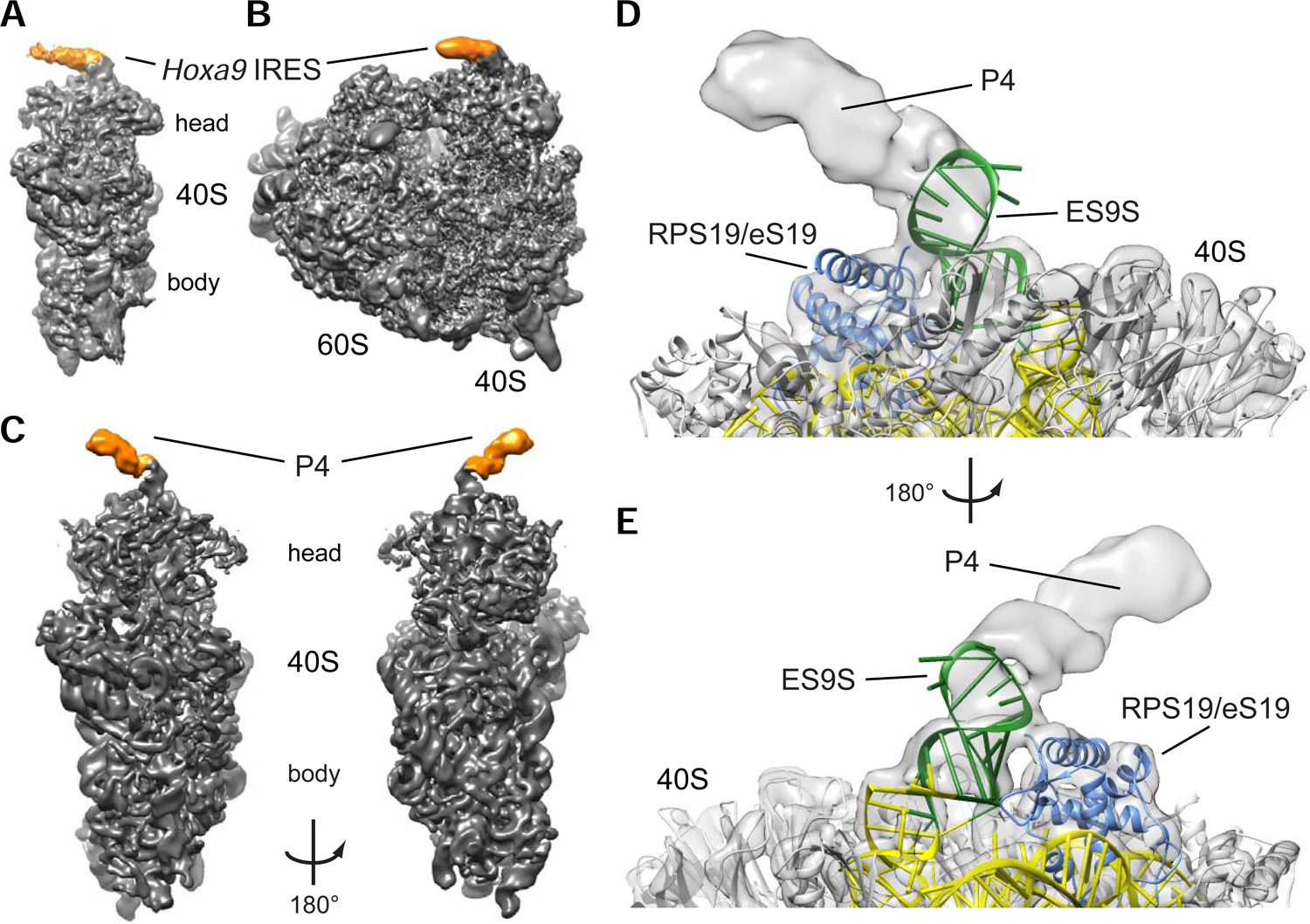
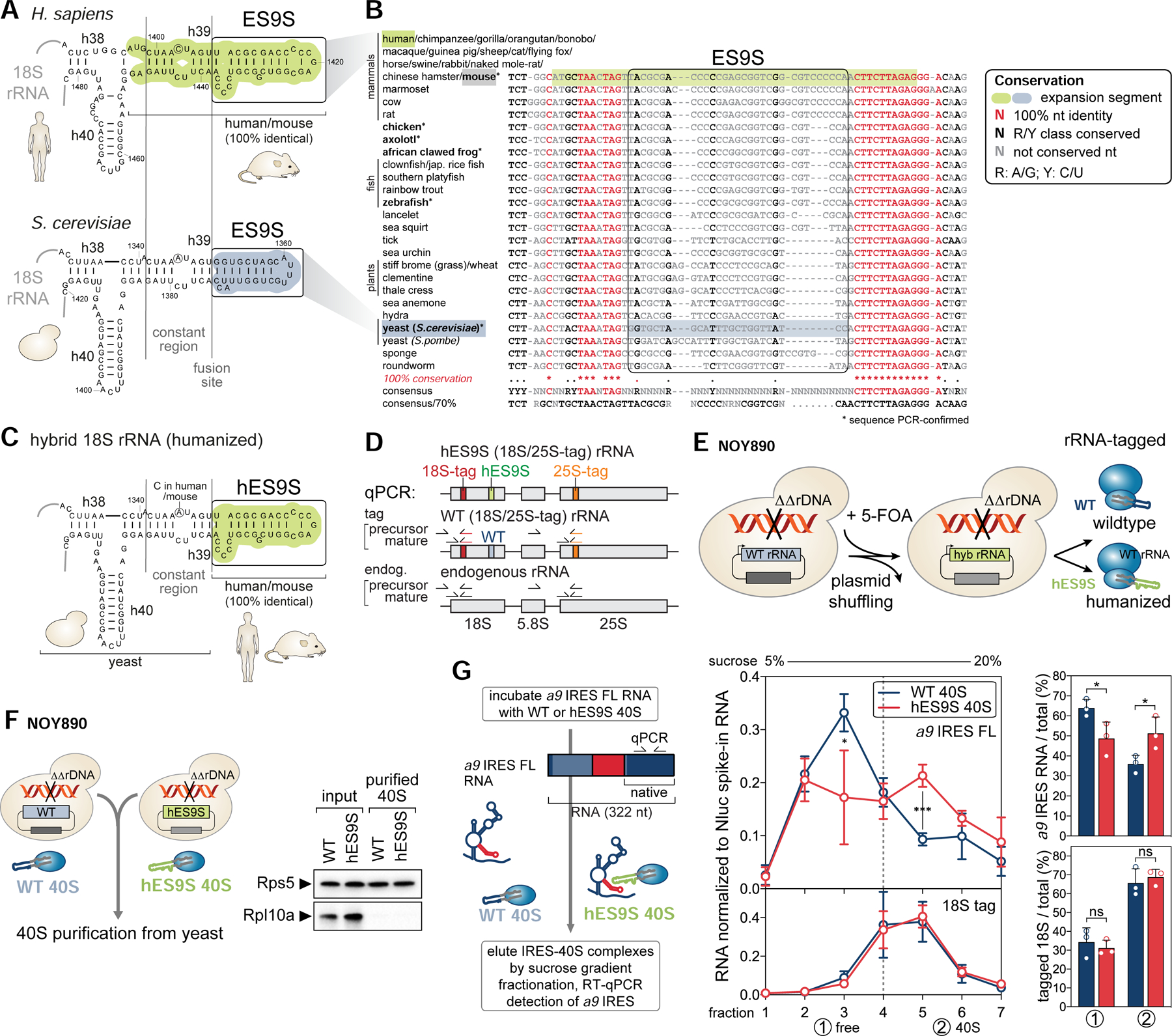
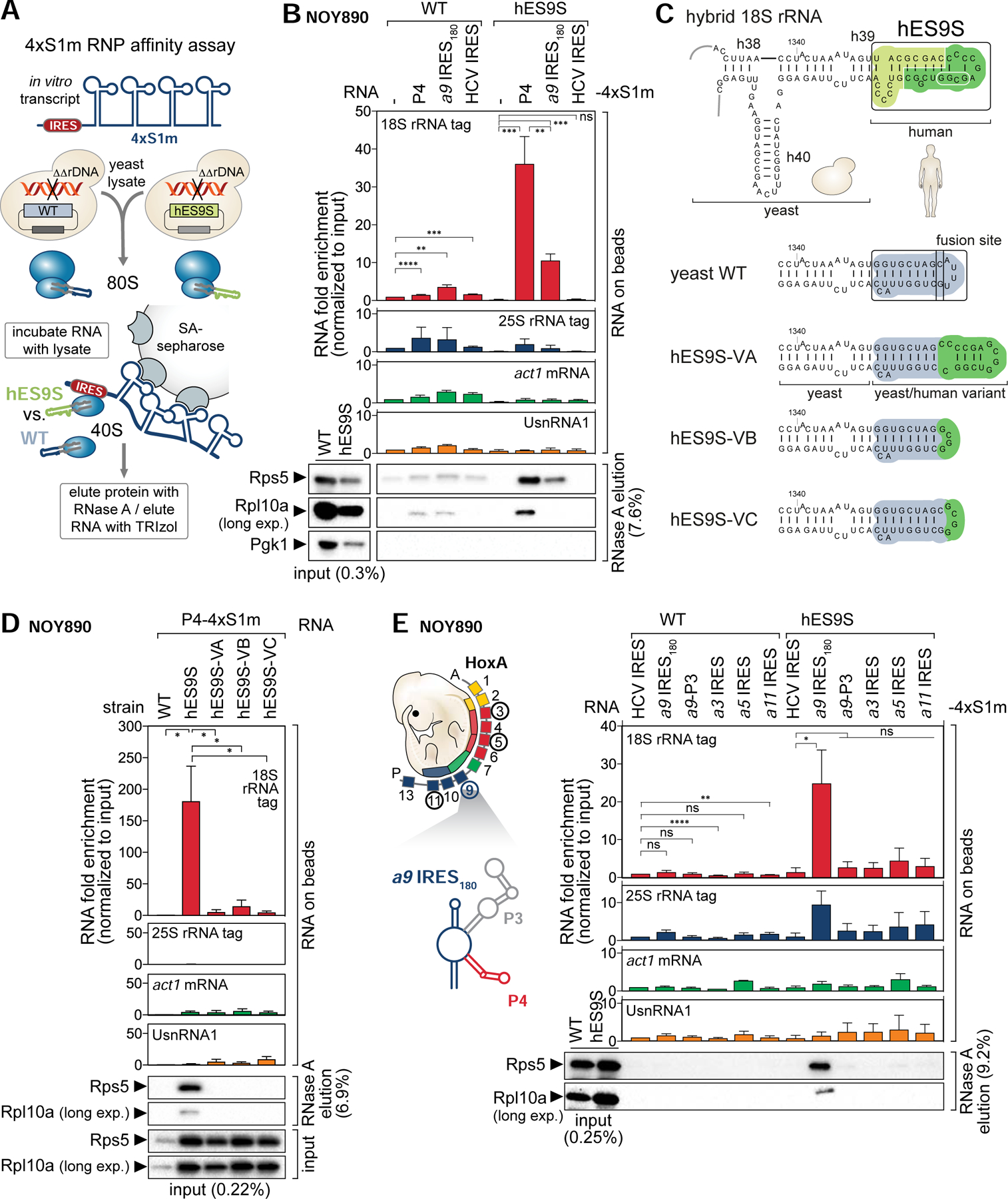
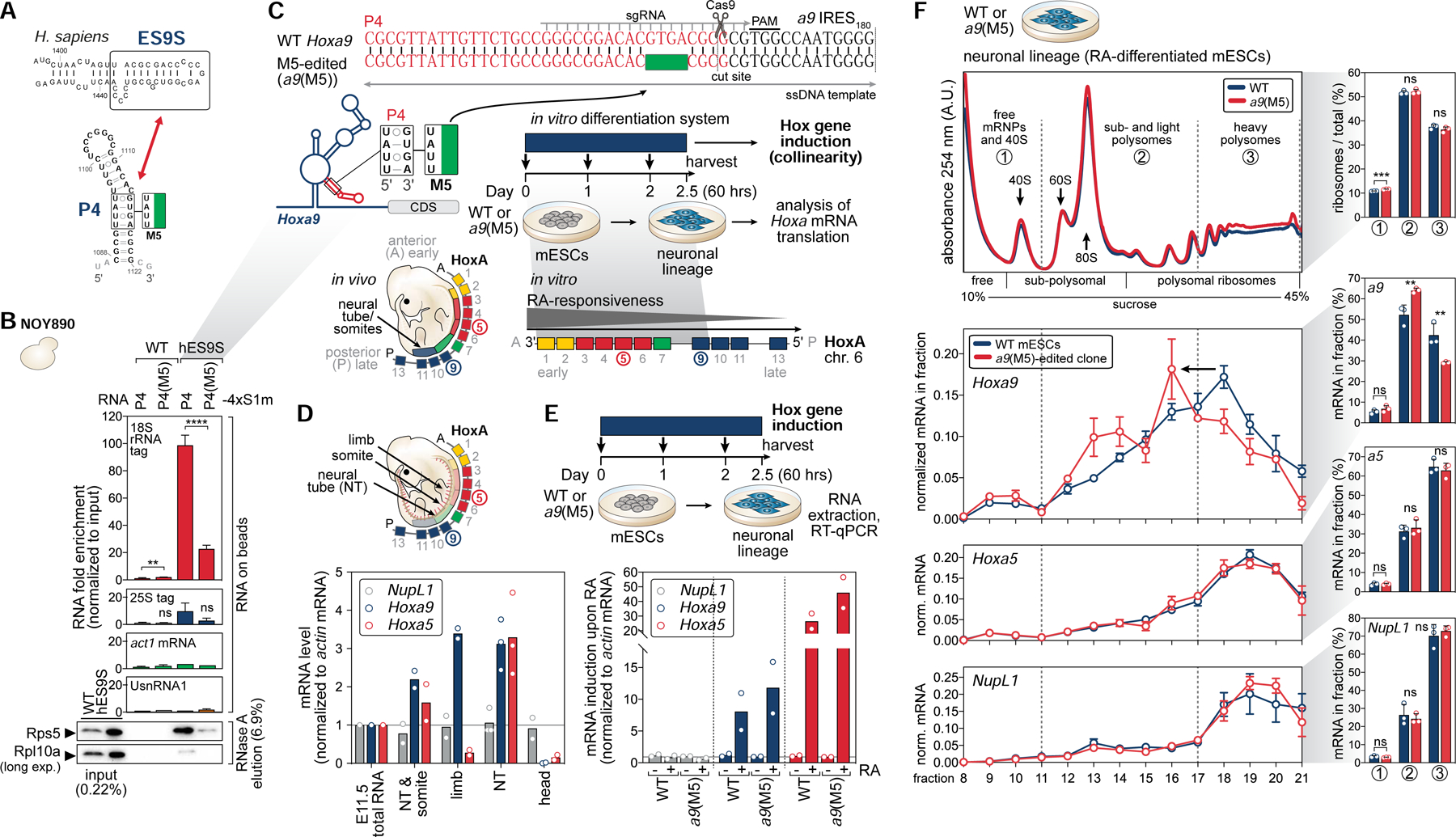
Ribosomes have been suggested to directly control gene regulation, but regulatory roles for ribosomal RNA (rRNA) remain largely unexplored. Expansion segments (ESs) consist of multitudes of tentacle-like rRNA structures extending from the core ribosome in eukaryotes. ESs are remarkably variable in sequence and size across eukaryotic evolution with largely unknown functions. In characterizing ribosome binding to a regulatory element within a Homeobox (Hox) 5' UTR, we identify a modular stem-loop within this element that binds to a single ES, ES9S. Engineering chimeric, "humanized" yeast ribosomes for ES9S reveals that an evolutionary change in the sequence of ES9S endows species-specific binding of Hoxa9 mRNA to the ribosome. Genome editing to site-specifically disrupt the Hoxa9-ES9S interaction demonstrates the functional importance for such selective mRNA-rRNA binding in translation control. Together, these studies unravel unexpected gene regulation directly mediated by rRNA and how ribosome evolution drives translation of critical developmental regulators.
Keywords: ES9S; Hox cluster; RNA structure; RNA-protein interaction; expansion segment; internal initiation; internal ribosome entry site; mRNA translation; ribosomal RNA; ribosome; ribosome engineering; stem-loop; yeast.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interest K.L. and M.B. are inventors on patents and submitted provisional patent applications related to the Hoxa9 P4 stem-loop and RNA therapeutics and their various uses.
Figures
References
-
- Acker MG, Kolitz SE, Mitchell SF, Nanda JS, and Lorsch JR (2007). Reconstitution of Yeast Translation Initiation. In Methods in Enzymology, pp. 111–145. - PubMed
-
- Anger AM, Armache JP, Berninghausen O, Habeck M, Subklewe M, Wilson DN, and Beckmann R (2013). Structures of the human and Drosophila 80S ribosome. Nature 497, 80–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
