

Navigating the crowd: visualizing coordination between genome dynamics, structure, and transcription
- PMID: 33203432
- PMCID: PMC7670612
- DOI: 10.1186/s13059-020-02185-y
Navigating the crowd: visualizing coordination between genome dynamics, structure, and transcription
Abstract
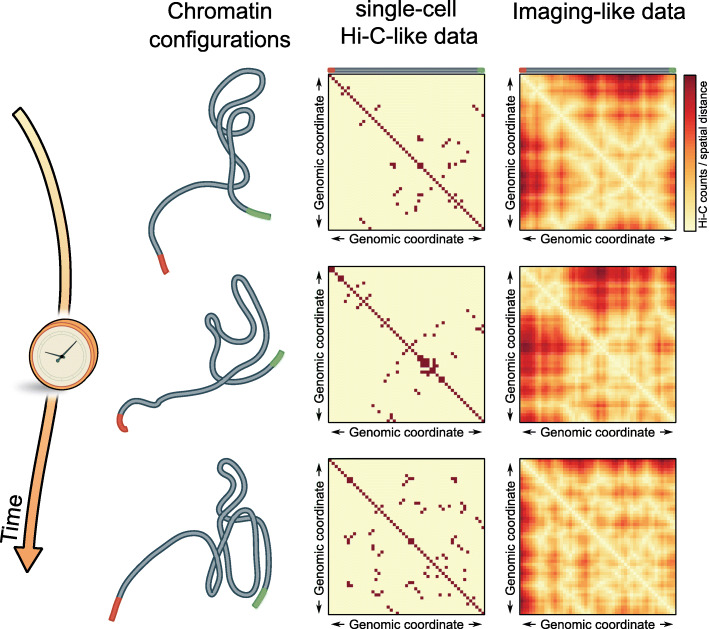
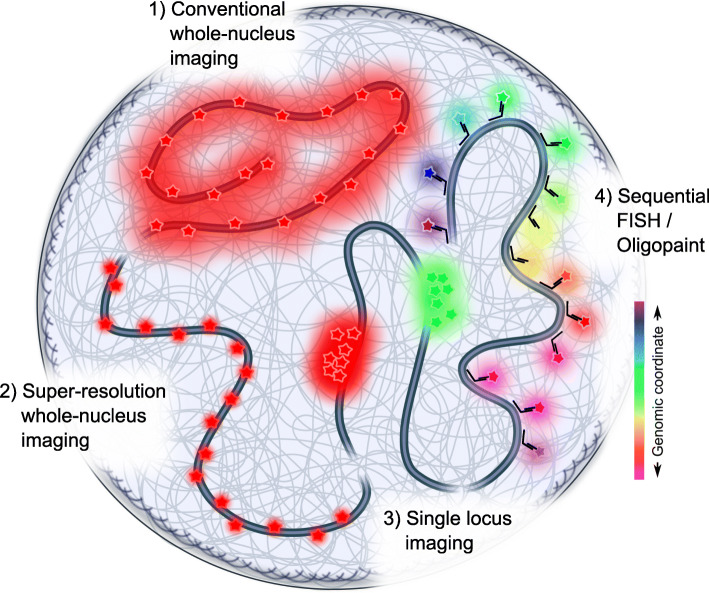
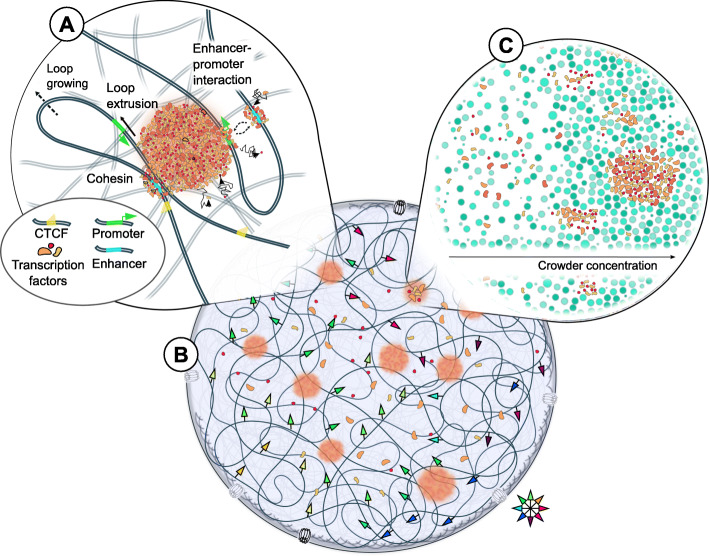
The eukaryotic genome is hierarchically structured yet highly dynamic. Regulating transcription in this environment demands a high level of coordination to permit many proteins to interact with chromatin fiber at appropriate sites in a timely manner. We describe how recent advances in quantitative imaging techniques overcome caveats of sequencing-based methods (Hi-C and related) by enabling direct visualization of transcription factors and chromatin at high resolution, from single genes to the whole nucleus. We discuss the contribution of fluorescence imaging to deciphering the principles underlying this coordination within the crowded nuclear space in living cells and discuss challenges ahead.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Transcription dynamics and genome organization in the mammalian nucleus: Recent advances.Mol Cell. 2025 Jan 16;85(2):208-224. doi: 10.1016/j.molcel.2024.09.022. Epub 2024 Oct 15. Mol Cell. 2025. PMID: 39413793 Review.
-
Dynamic organization of transcription compartments is dependent on functional nuclear architecture.Biophys J. 2012 Sep 5;103(5):851-9. doi: 10.1016/j.bpj.2012.06.036. Biophys J. 2012. PMID: 23009834 Free PMC article.
-
Visualizing chromatin dynamics in intact cells.Biochim Biophys Acta. 2008 Nov;1783(11):2044-51. doi: 10.1016/j.bbamcr.2008.06.022. Epub 2008 Jul 16. Biochim Biophys Acta. 2008. PMID: 18675855 Free PMC article. Review.
-
Hi-D: nanoscale mapping of nuclear dynamics in single living cells.Genome Biol. 2020 Apr 20;21(1):95. doi: 10.1186/s13059-020-02002-6. Genome Biol. 2020. PMID: 32312289 Free PMC article.
-
Geometric control and modeling of genome reprogramming.Bioarchitecture. 2016 Jul 3;6(4):76-84. doi: 10.1080/19490992.2016.1201620. Epub 2016 Jul 19. Bioarchitecture. 2016. PMID: 27434579 Free PMC article.
Cited by
-
Chromatin loops gather targets of upstream regulators together for efficient gene transcription regulation during vernalization in wheat.Genome Biol. 2024 Dec 3;25(1):306. doi: 10.1186/s13059-024-03437-x. Genome Biol. 2024. PMID: 39623466 Free PMC article.
-
Tracing the Chromatin: From 3C to Live-Cell Imaging.Chem Biomed Imaging. 2024 Jun 25;2(10):659-682. doi: 10.1021/cbmi.4c00033. eCollection 2024 Oct 28. Chem Biomed Imaging. 2024. PMID: 39483638 Free PMC article. Review.
-
Chromatin Network Analyses: Towards Structure-Function Relationships in Epigenomics.Front Bioinform. 2021 Oct 27;1:742216. doi: 10.3389/fbinf.2021.742216. eCollection 2021. Front Bioinform. 2021. PMID: 36303769 Free PMC article.
-
Human RNA Polymerase II Segregates from Genes and Nascent RNA and Transcribes in the Presence of DNA-Bound dCas9.Int J Mol Sci. 2024 Aug 1;25(15):8411. doi: 10.3390/ijms25158411. Int J Mol Sci. 2024. PMID: 39125980 Free PMC article.
-
The anatomy of transcriptionally active chromatin loops in Drosophila primary spermatocytes using super-resolution microscopy.PLoS Genet. 2023 Mar 3;19(3):e1010654. doi: 10.1371/journal.pgen.1010654. eCollection 2023 Mar. PLoS Genet. 2023. PMID: 36867662 Free PMC article.
References
-
- Lanctôt C, Cheutin T, Cremer M, Cavalli G, Cremer T. Dynamic genome architecture in the nuclear space: regulation of gene expression in three dimensions. Nat Rev Genet. 2007;8(2):104-15. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
