

Consistent population declines but idiosyncratic range shifts in Alpine orchids under global change
- PMID: 33203870
- PMCID: PMC7672077
- DOI: 10.1038/s41467-020-19680-2
Consistent population declines but idiosyncratic range shifts in Alpine orchids under global change
Abstract
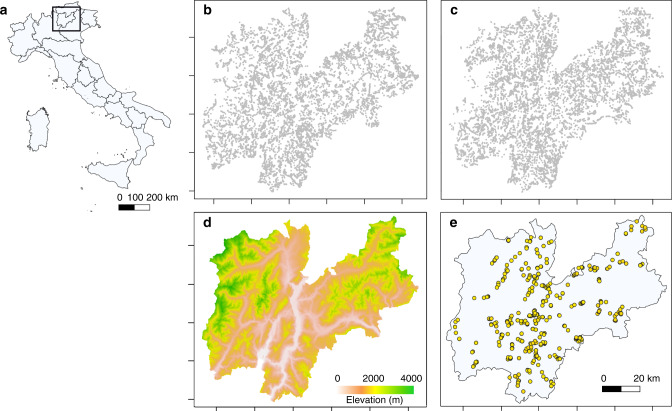
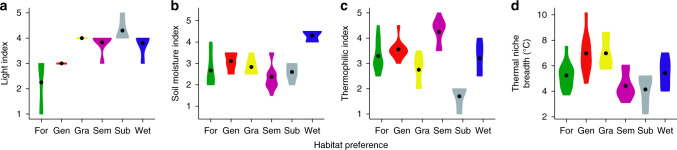
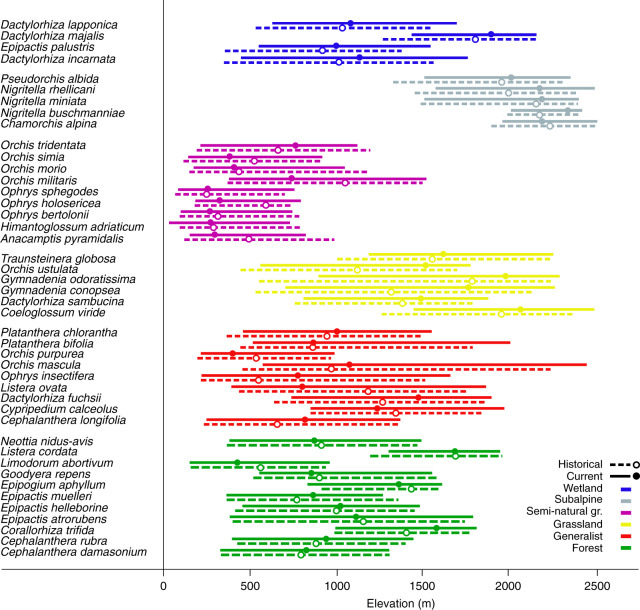
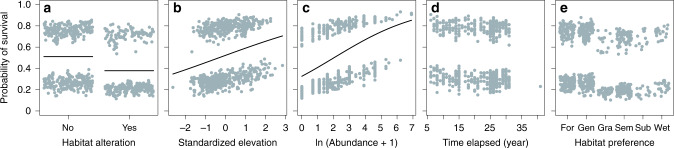
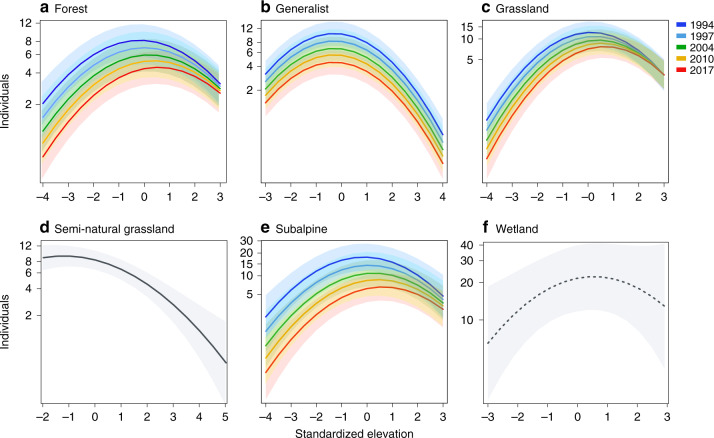
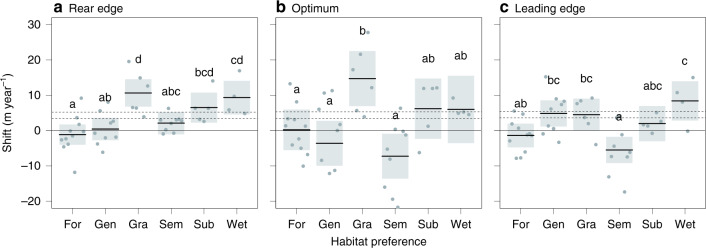
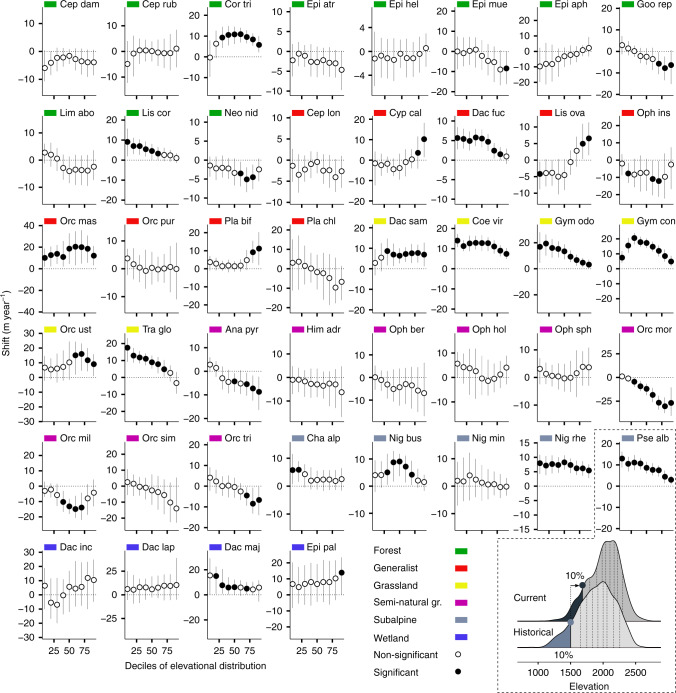
Mountains are plant biodiversity hotspots considered particularly vulnerable to multiple environmental changes. Here, we quantify population changes and range-shift dynamics along elevational gradients over the last three decades for c. two-thirds of the orchid species of the European Alps. Local extinctions were more likely for small populations, after habitat alteration, and predominated at the rear edge of species' ranges. Except for the most thermophilic species and wetland specialists, population density decreased over time. Declines were more pronounced for rear-edge populations, possibly due to multiple pressures such as climate warming, habitat alteration, and mismatched ecological interactions. Besides these demographic trends, different species exhibited idiosyncratic range shifts with more than 50% of the species lagging behind climate warming. Our study highlights the importance of long-term monitoring of populations and range distributions at fine spatial resolution to be able to fully understand the consequences of global change for orchids.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Gottfried M, et al. Continent-wide response of mountain vegetation to climate change. Nat. Clim. Chang. 2012;2:111–115. doi: 10.1038/nclimate1329. - DOI
-
- Dainese M, et al. Human disturbance and upward expansion of plants in a warming climate. Nat. Clim. Chang. 2017;7:577–580. doi: 10.1038/nclimate3337. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
