

TRAF2 regulates T cell immunity by maintaining a Tpl2-ERK survival signaling axis in effector and memory CD8 T cells
- PMID: 33203937
- PMCID: PMC8429472
- DOI: 10.1038/s41423-020-00583-7
TRAF2 regulates T cell immunity by maintaining a Tpl2-ERK survival signaling axis in effector and memory CD8 T cells
Abstract
Generation and maintenance of antigen-specific effector and memory T cells are central events in immune responses against infections. We show that TNF receptor-associated factor 2 (TRAF2) maintains a survival signaling axis in effector and memory CD8 T cells required for immune responses against infections. This signaling axis involves activation of Tpl2 and its downstream kinase ERK by NF-κB-inducing kinase (NIK) and degradation of the proapoptotic factor Bim. NIK mediates Tpl2 activation by stimulating the phosphorylation and degradation of the Tpl2 inhibitor p105. Interestingly, while NIK is required for Tpl2-ERK signaling under normal conditions, uncontrolled NIK activation due to loss of its negative regulator, TRAF2, causes constitutive degradation of p105 and Tpl2, leading to severe defects in ERK activation and effector/memory CD8 T cell survival. Thus, TRAF2 controls a previously unappreciated signaling axis mediating effector/memory CD8 T cell survival and protective immunity.
Keywords: Bacterial infection; Effector and memory CD8 T cells; NIK; Protective immunity; T cell survival; TRAF2; Tpl2.
© 2020. CSI and USTC.
Conflict of interest statement
The authors declare no competing interests.
Figures
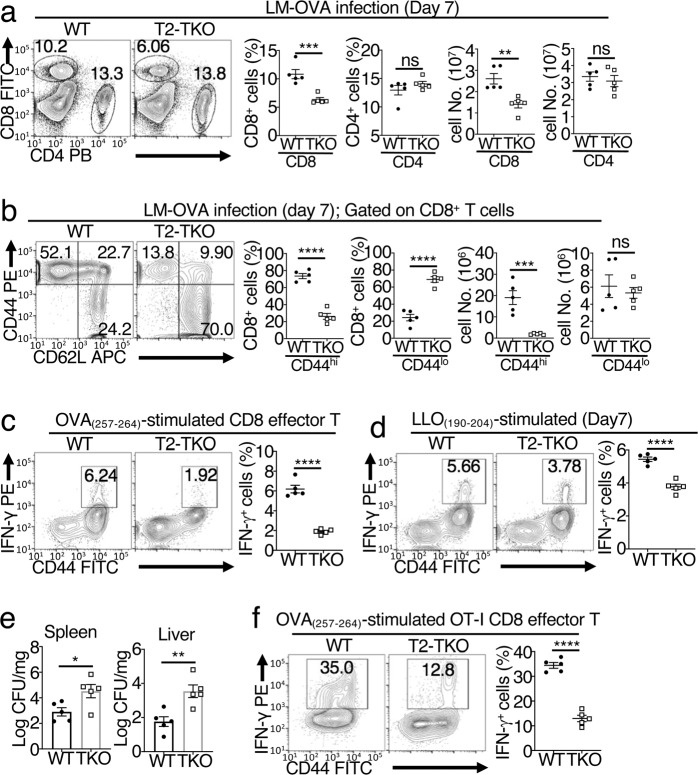
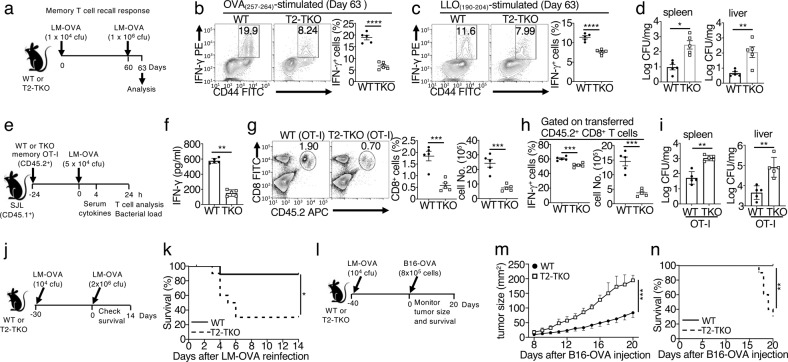
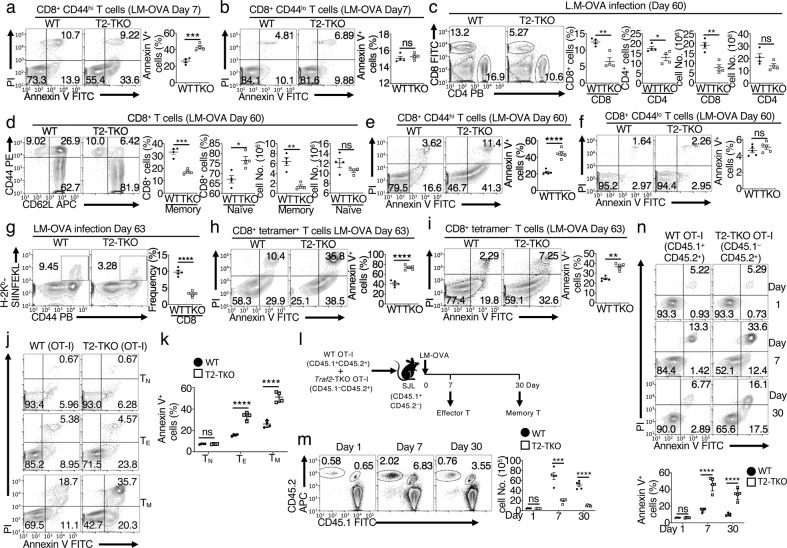
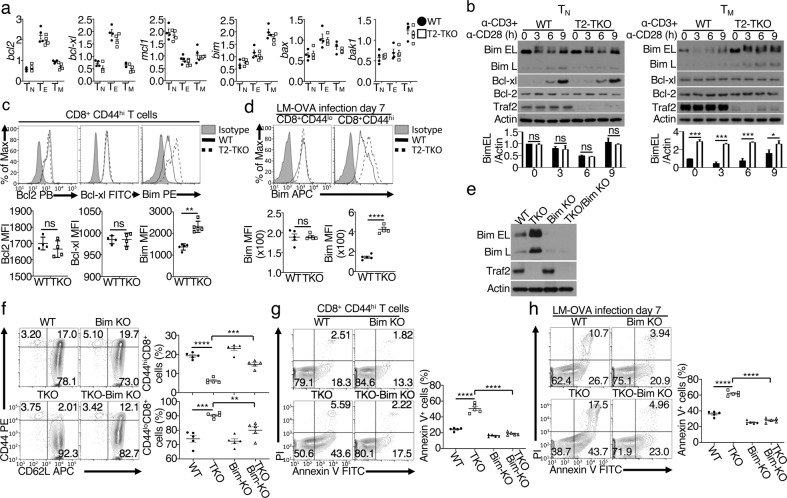
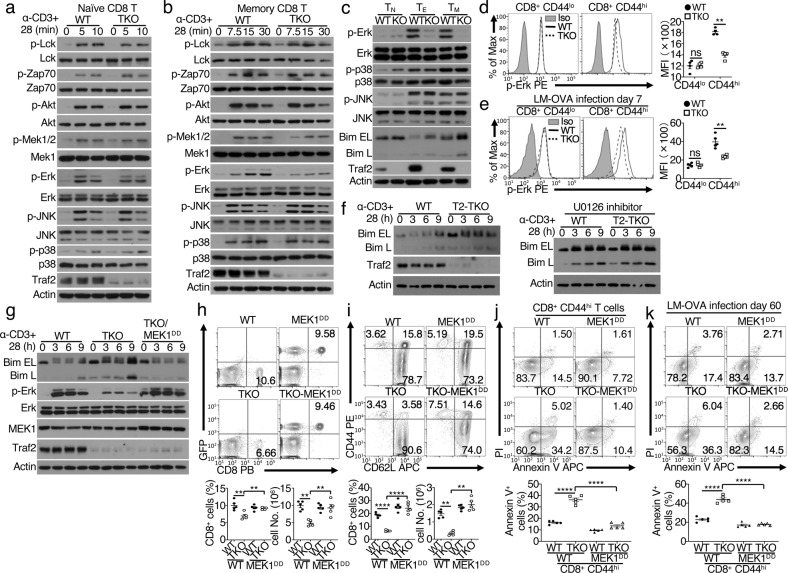
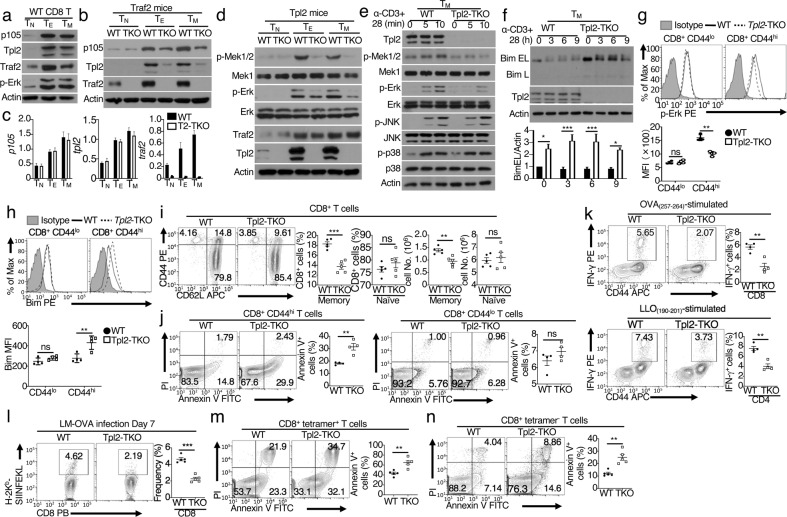
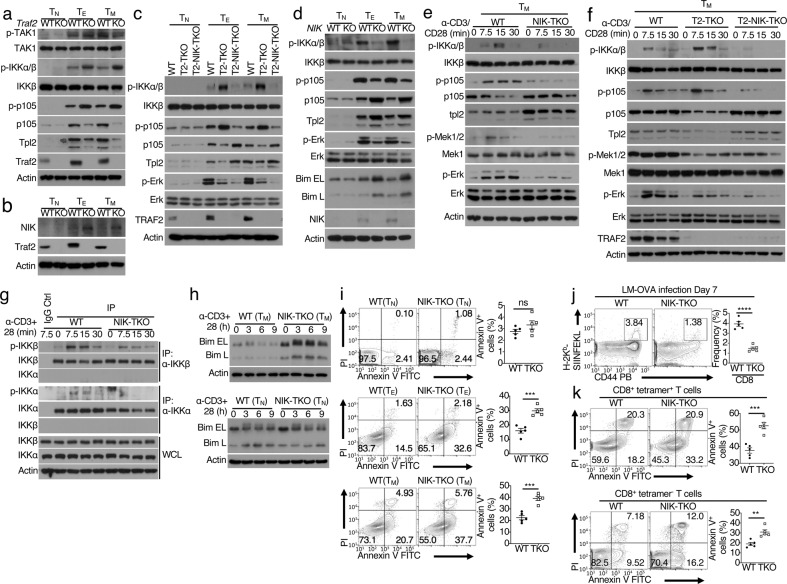
References
-
- Glimcher LH, Townsend MJ, Sullivan BM, Lord GM. Recent developments in the transcriptional regulation of cytolytic effector cells. Nat. Rev. Immunol. 2004;4:900–911. - PubMed
-
- Marsden VS, Strasser A. Control of apoptosis in the immune system: Bcl-2, BH3-only proteins and more. Annu. Rev. Immunol. 2003;21:71–105. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
