

Dendritic and parallel processing of visual threats in the retina control defensive responses
- PMID: 33208370
- PMCID: PMC7673819
- DOI: 10.1126/sciadv.abc9920
Dendritic and parallel processing of visual threats in the retina control defensive responses
Abstract
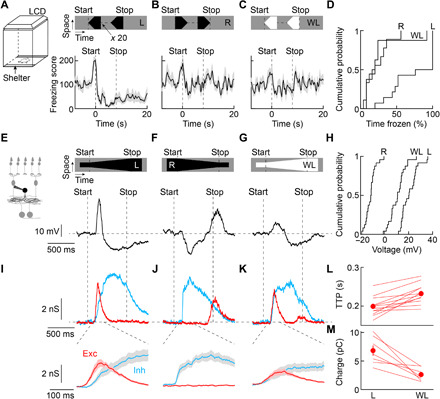
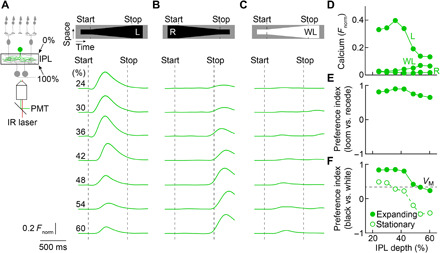
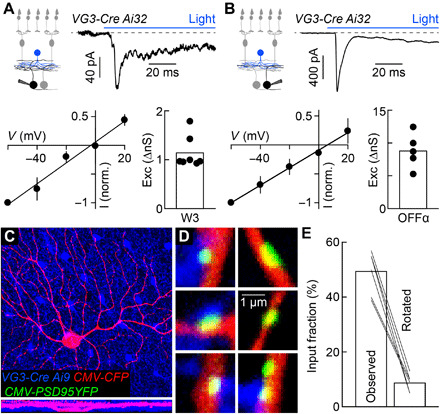
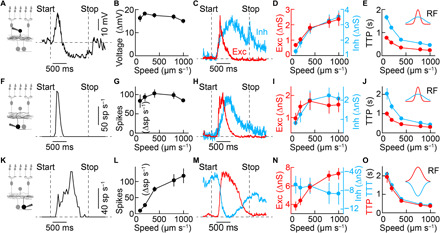
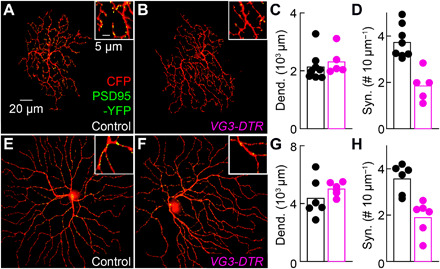
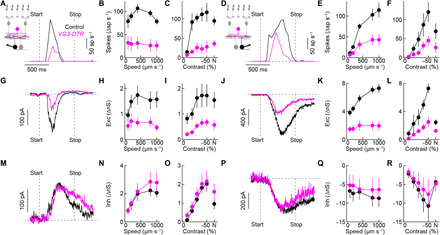
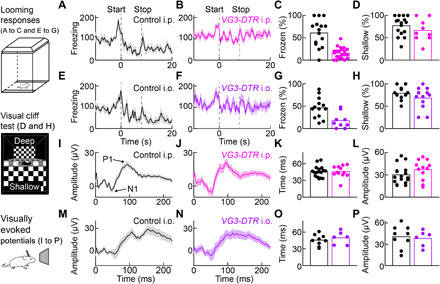
Approaching predators cast expanding shadows (i.e., looming) that elicit innate defensive responses in most animals. Where looming is first detected and how critical parameters of predatory approaches are extracted are unclear. In mice, we identify a retinal interneuron (the VG3 amacrine cell) that responds robustly to looming, but not to related forms of motion. Looming-sensitive calcium transients are restricted to a specific layer of the VG3 dendrite arbor, which provides glutamatergic input to two ganglion cells (W3 and OFFα). These projection neurons combine shared excitation with dissimilar inhibition to signal approach onset and speed, respectively. Removal of VG3 amacrine cells reduces the excitation of W3 and OFFα ganglion cells and diminishes defensive responses of mice to looming without affecting other visual behaviors. Thus, the dendrites of a retinal interneuron detect visual threats, divergent circuits downstream extract critical threat parameters, and these retinal computations initiate an innate survival behavior.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Fotowat H., Gabbiani F., Collision detection as a model for sensory-motor integration. Annu. Rev. Neurosci. 34, 1–19 (2011). - PubMed
-
- Peek M. Y., Card G. M., Comparative approaches to escape. Curr. Opin. Neurobiol. 41, 167–173 (2016). - PubMed
-
- De Franceschi G., Vivattanasarn T., Saleem A. B., Solomon S. G., Vision guides selection of freeze or flight defense strategies in mice. Curr. Biol. 26, 2150–2154 (2016). - PubMed
-
- Shang C., Liu Z., Chen Z., Shi Y., Wang Q., Liu S., Li D., Cao P., A parvalbumin-positive excitatory visual pathway to trigger fear responses in mice. Science 348, 1472–1477 (2015). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
