

Golgi-Dependent Copper Homeostasis Sustains Synaptic Development and Mitochondrial Content
- PMID: 33208468
- PMCID: PMC7810662
- DOI: 10.1523/JNEUROSCI.1284-20.2020
Golgi-Dependent Copper Homeostasis Sustains Synaptic Development and Mitochondrial Content
Abstract
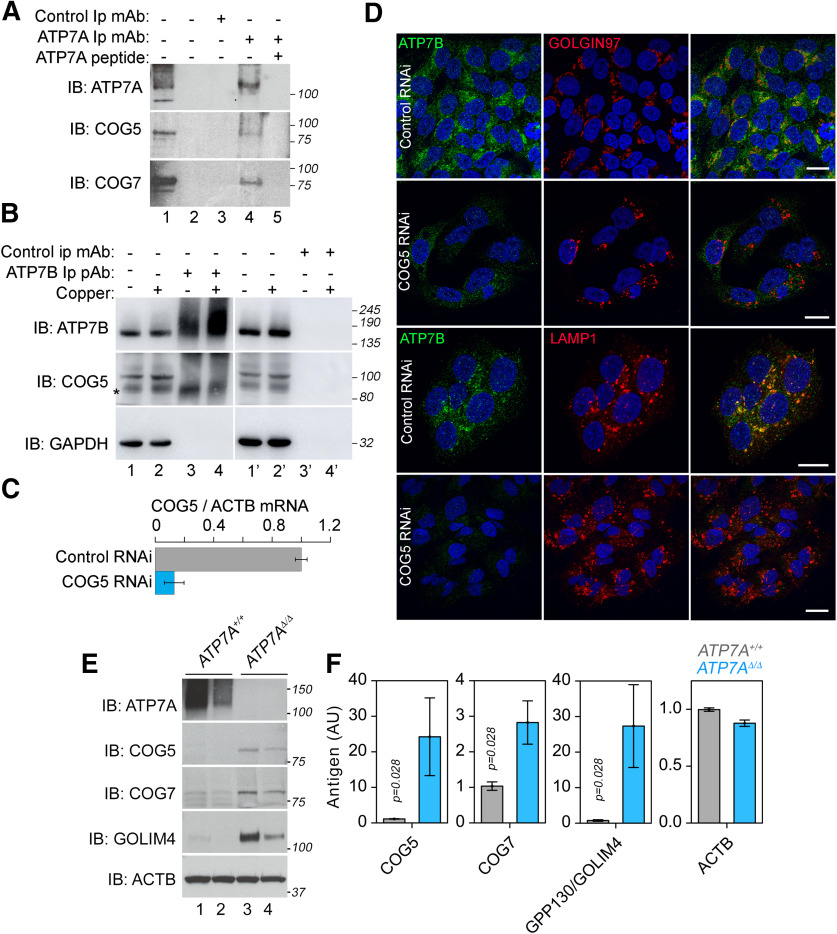
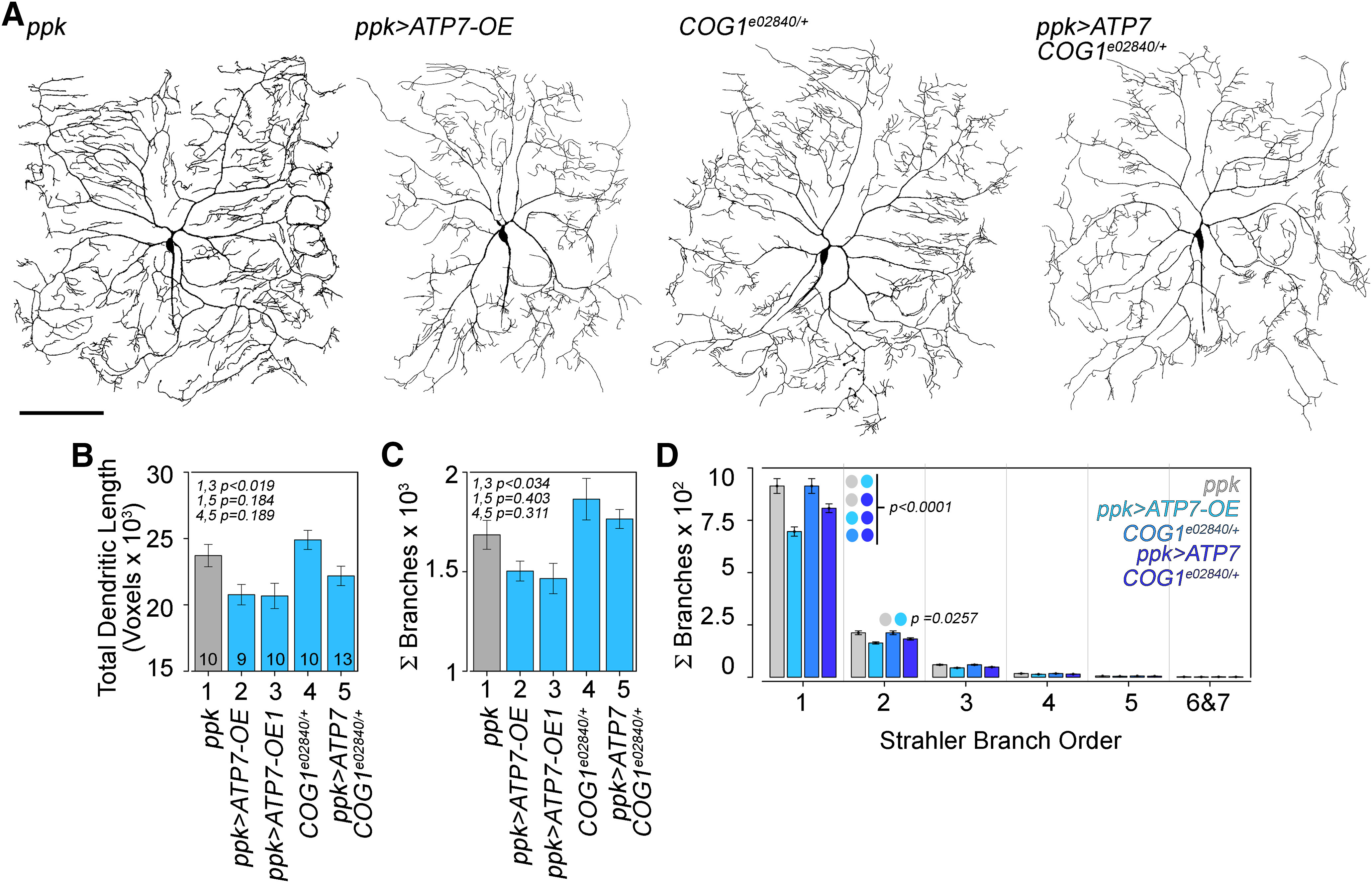
Rare genetic diseases preponderantly affect the nervous system causing neurodegeneration to neurodevelopmental disorders. This is the case for both Menkes and Wilson disease, arising from mutations in ATP7A and ATP7B, respectively. The ATP7A and ATP7B proteins localize to the Golgi and regulate copper homeostasis. We demonstrate genetic and biochemical interactions between ATP7 paralogs with the conserved oligomeric Golgi (COG) complex, a Golgi apparatus vesicular tether. Disruption of Drosophila copper homeostasis by ATP7 tissue-specific transgenic expression caused alterations in epidermis, aminergic, sensory, and motor neurons. Prominent among neuronal phenotypes was a decreased mitochondrial content at synapses, a phenotype that paralleled with alterations of synaptic morphology, transmission, and plasticity. These neuronal and synaptic phenotypes caused by transgenic expression of ATP7 were rescued by downregulation of COG complex subunits. We conclude that the integrity of Golgi-dependent copper homeostasis mechanisms, requiring ATP7 and COG, are necessary to maintain mitochondria functional integrity and localization to synapses.SIGNIFICANCE STATEMENT Menkes and Wilson disease affect copper homeostasis and characteristically afflict the nervous system. However, their molecular neuropathology mechanisms remain mostly unexplored. We demonstrate that copper homeostasis in neurons is maintained by two factors that localize to the Golgi apparatus, ATP7 and the conserved oligomeric Golgi (COG) complex. Disruption of these mechanisms affect mitochondrial function and localization to synapses as well as neurotransmission and synaptic plasticity. These findings suggest communication between the Golgi apparatus and mitochondria through homeostatically controlled cellular copper levels and copper-dependent enzymatic activities in both organelles.
Keywords: ATP7A; Golgi; Menkes; Wilson; copper; mitochondria.
Copyright © 2021 the authors.
Figures








References
-
- Allensworth JL, Evans MK, Bertucci F, Aldrich AJ, Festa RA, Finetti P, Ueno NT, Safi R, McDonnell DP, Thiele DJ, Van Laere S, Devi GR (2015) Disulfiram (DSF) acts as a copper ionophore to induce copper-dependent oxidative stress and mediate anti-tumor efficacy in inflammatory breast cancer. Mol Oncol 9:1155–1168. 10.1016/j.molonc.2015.02.007 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases