

Persistently activated, proliferative memory autoreactive B cells promote inflammation in rheumatoid arthritis
- PMID: 33208502
- PMCID: PMC7615909
- DOI: 10.1126/scitranslmed.aaz5327
Persistently activated, proliferative memory autoreactive B cells promote inflammation in rheumatoid arthritis
Abstract
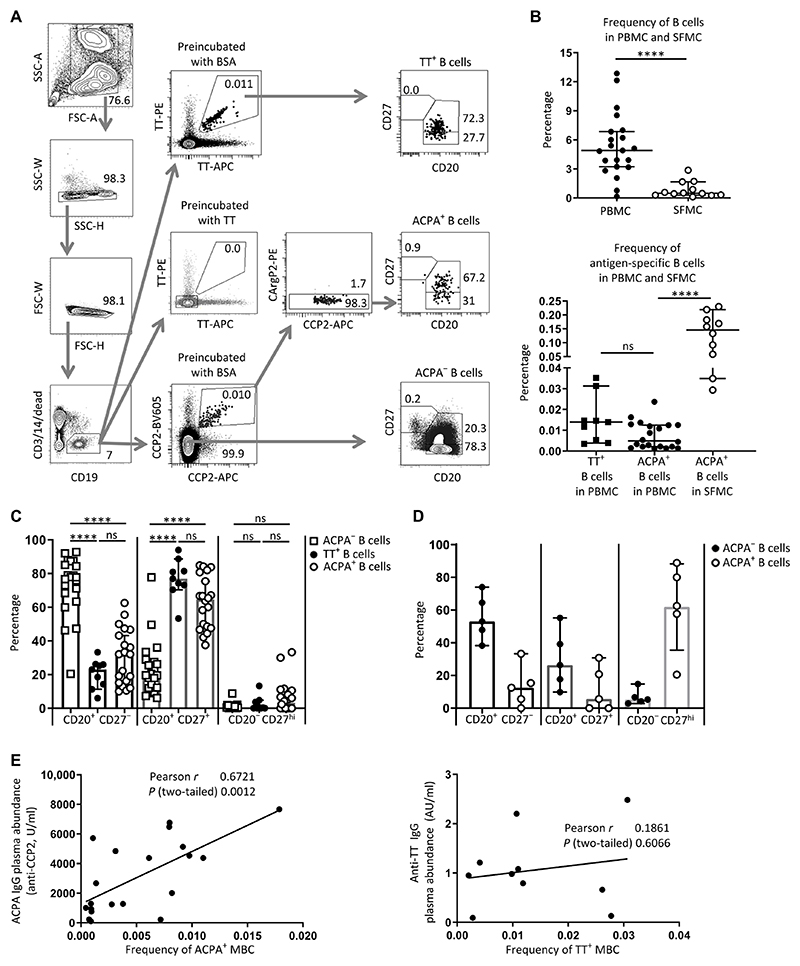
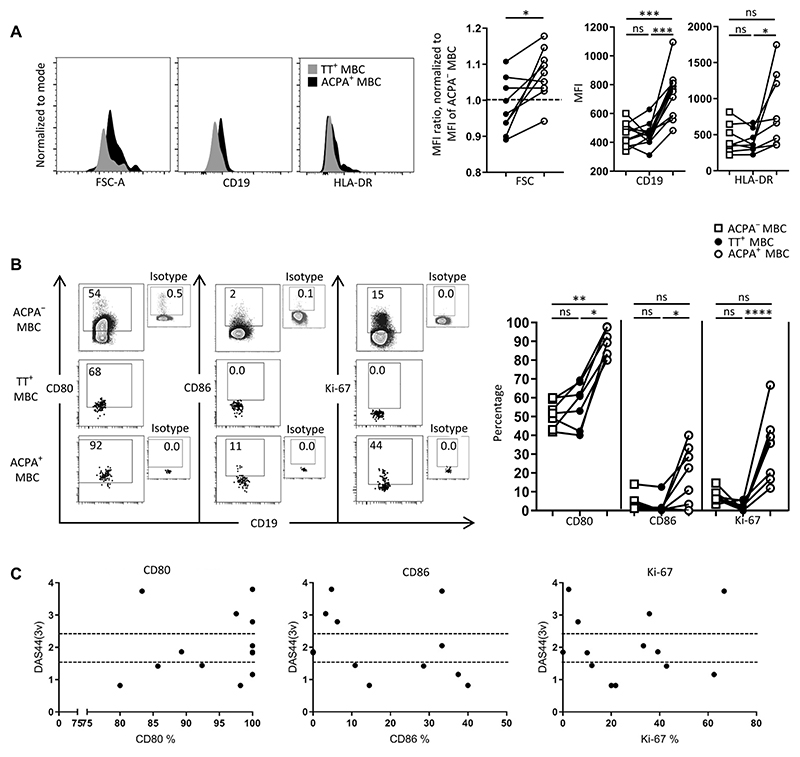
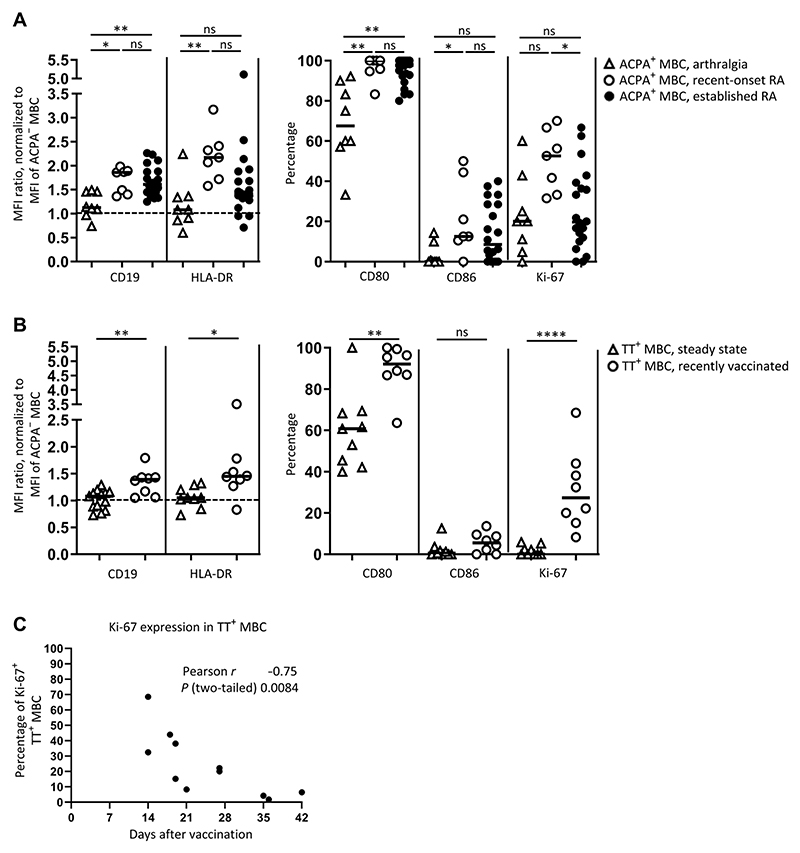
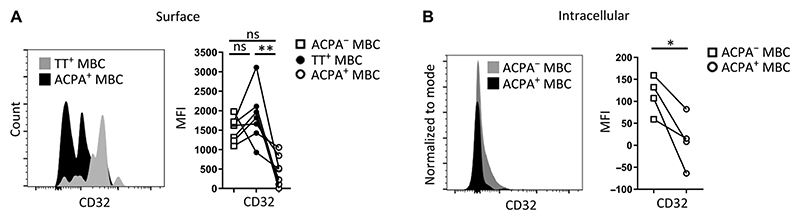
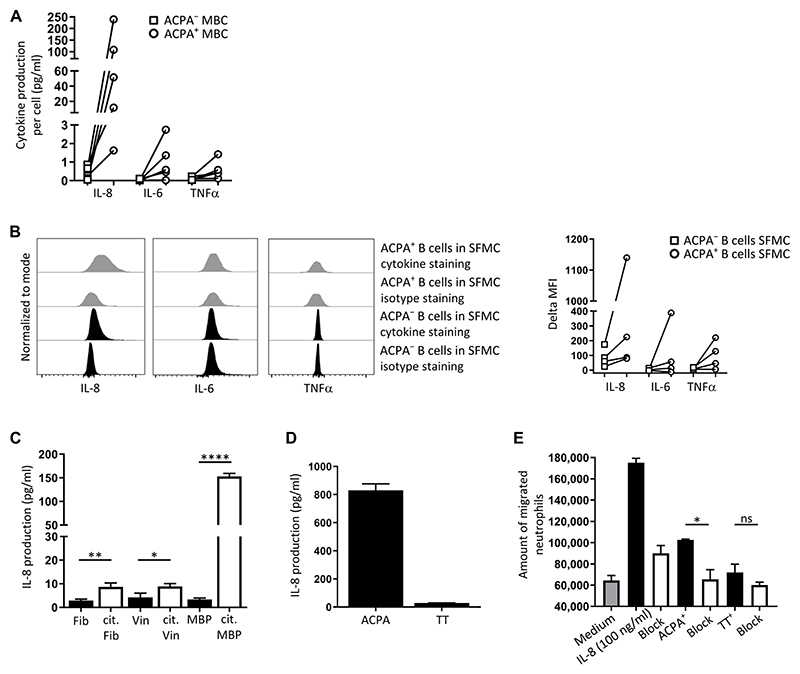
Autoreactive B cells mediate autoimmune pathology, but exactly how remains unknown. A hallmark of rheumatoid arthritis (RA), a common autoimmune disease, is the presence of disease-specific anticitrullinated protein antibodies (ACPAs). Here, we showed that ACPA-positive B cells in patients with RA strongly expressed T cell-stimulating ligands, produced abundant proinflammatory cytokines, and were proliferative while escaping inhibitory signals. This activated state was found at different degrees in different stages of disease: highest in patients with recent-onset RA, moderate in patients with established RA, and far less pronounced in ACPA-positive individuals "at risk" for developing disease. The activated autoreactive B cell response persisted in patients who achieved clinical remission with conventional treatment. ACPA-positive B cells in blood and synovial fluid secreted increased amounts of the chemoattractant interleukin-8, which attracted neutrophils, the most abundant immune cell in arthritic joints. Tetanus toxoid-specific B cells from the same patients exhibited properties of memory B cells without the activation and proliferation phenotype, but these cells transiently acquired a similar proliferative phenotype upon booster vaccination. Together, these data indicated that continuous antigenic triggering of autoreactive B cells occurs in human autoimmune disease and support the emerging concept of immunological activity that persists under treatment even in clinical remission, which may revise our current concept of treatment targets for future therapeutic interventions. In addition, our data pointed to a pathogenic role of ACPA-positive B cells in the inflammatory disease process underlying RA and favor approaches that aim at their antigen-specific inactivation or depletion.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Scherer HU, Huizinga TWJ, Krönke G, Schett G, Toes REM. The B cell response to citrullinated antigens in the development of rheumatoid arthritis. Nat Rev Rheumatol. 2018;14:157–169. - PubMed
-
- Willemze A, Trouw LA, Toes RE, Huizinga TW. The influence of ACPA status and characteristics on the course of RA. Nat Rev Rheumatol. 2012;8:144–152. - PubMed
-
- Burgers LE, van Steenbergen HW, Ten Brinck RM, Huizinga TW, van der Helm-van Mil AH. Differences in the symptomatic phase preceding ACPA-positive and ACPA-negative RA: A longitudinal study in arthralgia during progression to clinical arthritis. Ann Rheum Dis. 2017;76:1751–1754. - PubMed
-
- van der Woude D, Rantapää-Dahlqvist S, Ioan-Facsinay A, Onnekink C, Schwarte CM, Verpoort KN, Drijfhout JW, Huizinga TW, Toes RE, Pruijn GJ. Epitope spreading of the anti-citrullinated protein antibody response occurs before disease onset and is associated with the disease course of early arthritis. Ann Rheum Dis. 2010;69:1554–1561. - PubMed
-
- Suwannalai P, van de Stadt LA, Radner H, Steiner G, El-Gabalawy HS, Zijde CM, van Tol MJ, van Schaardenburg D, Huizinga TW, Toes RE, Trouw LA. Avidity maturation of anti-citrullinated protein antibodies in rheumatoid arthritis. Arthritis Rheum. 2012;64:1323–1328. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
