

Identification of Spacer and Protospacer Sequence Requirements in the Vibrio cholerae Type I-E CRISPR/Cas System
- PMID: 33208517
- PMCID: PMC7677007
- DOI: 10.1128/mSphere.00813-20
Identification of Spacer and Protospacer Sequence Requirements in the Vibrio cholerae Type I-E CRISPR/Cas System
Abstract
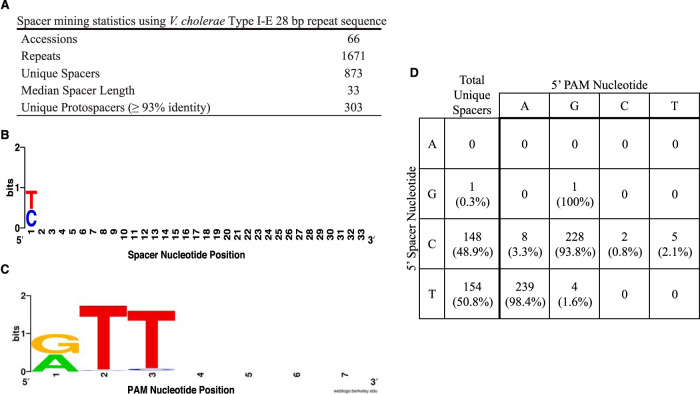
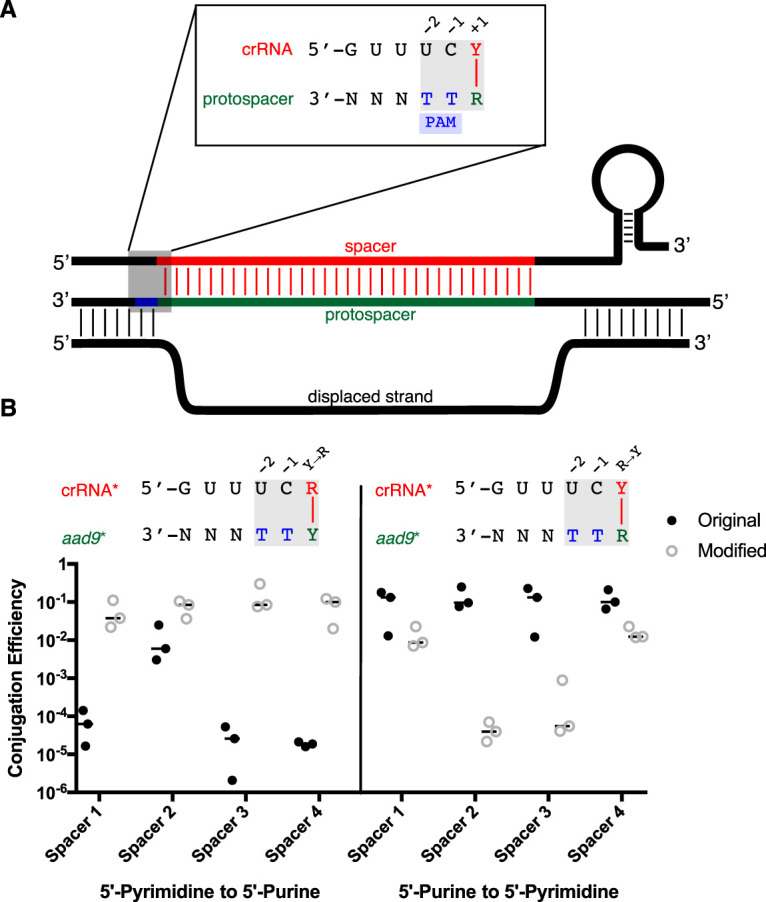
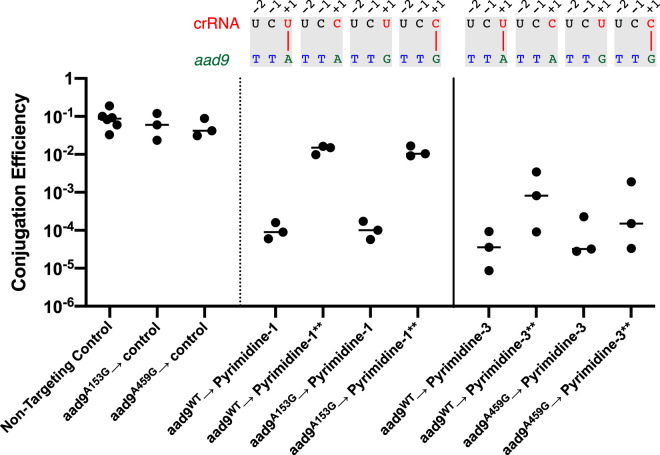
The prokaryotic adaptive immune system CRISPR/Cas serves as a defense against bacteriophage and invasive nucleic acids. A type I-E CRISPR/Cas system has been detected in classical biotype isolates of Vibrio cholerae, the causative agent of the disease cholera. Experimental characterization of this system revealed a functional immune system that operates using a 5'-TT-3' protospacer-adjacent motif (PAM) for interference. However, several designed spacers against the 5'-TT-3' PAM do not interfere as expected, indicating that further investigation of this system is necessary. In this study, we identified additional conserved sequences, including a pyrimidine in the 5' position of the spacer and a purine in the complementary position of the protospacer using 873 unique spacers and 2,267 protospacers mined from CRISPR arrays in deposited sequences of V. cholerae We present bioinformatic evidence that during acquisition the protospacer purine is captured in the prespacer and that a 5'-RTT-3' PAM is necessary for spacer acquisition. Finally, we demonstrate experimentally, by designing and manipulating spacer and cognate PAMs in a plasmid conjugation assay, that a 5'-RTT-3' PAM is necessary for CRISPR interference, and we discover functional consequences for spacer efficacy related to the identity of the 5' spacer pyrimidine.IMPORTANCE Bacterial CRISPR/Cas systems provide immunity by defending against phage and other invading elements. A thorough comprehension of the molecular mechanisms employed by these diverse systems will improve our understanding of bacteriophage-bacterium interactions and bacterial adaptation to foreign DNA. The Vibrio cholerae type I-E system was previously identified in an extinct classical biotype and was partially characterized for its function. Here, using both bioinformatic and functional assays, we extend that initial study. We have found that the type I-E system still exists in modern strains of V. cholerae Furthermore, we defined additional sequence elements both in the CRISPR array and in target DNA that are required for immunity. CRISPR/Cas systems are now commonly used as precise and powerful genetic engineering tools. Knowledge of the sequences required for CRISPR/Cas immunity is a prerequisite for the effective design and experimental use of these systems. Our results greatly facilitate the effective use of one such system. Furthermore, we provide a publicly available software program that assists in the detection and validation of CRISPR/Cas immunity requirements when such a system exists in a bacterial species.
Keywords: CRISPR/Cas; Vibrio cholerae; protospacer-adjacent motif.
Copyright © 2020 Bourgeois et al.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
