

Identification of the HECT E3 ligase UBR5 as a regulator of MYC degradation using a CRISPR/Cas9 screen
- PMID: 33208877
- PMCID: PMC7676242
- DOI: 10.1038/s41598-020-76960-z
Identification of the HECT E3 ligase UBR5 as a regulator of MYC degradation using a CRISPR/Cas9 screen
Abstract
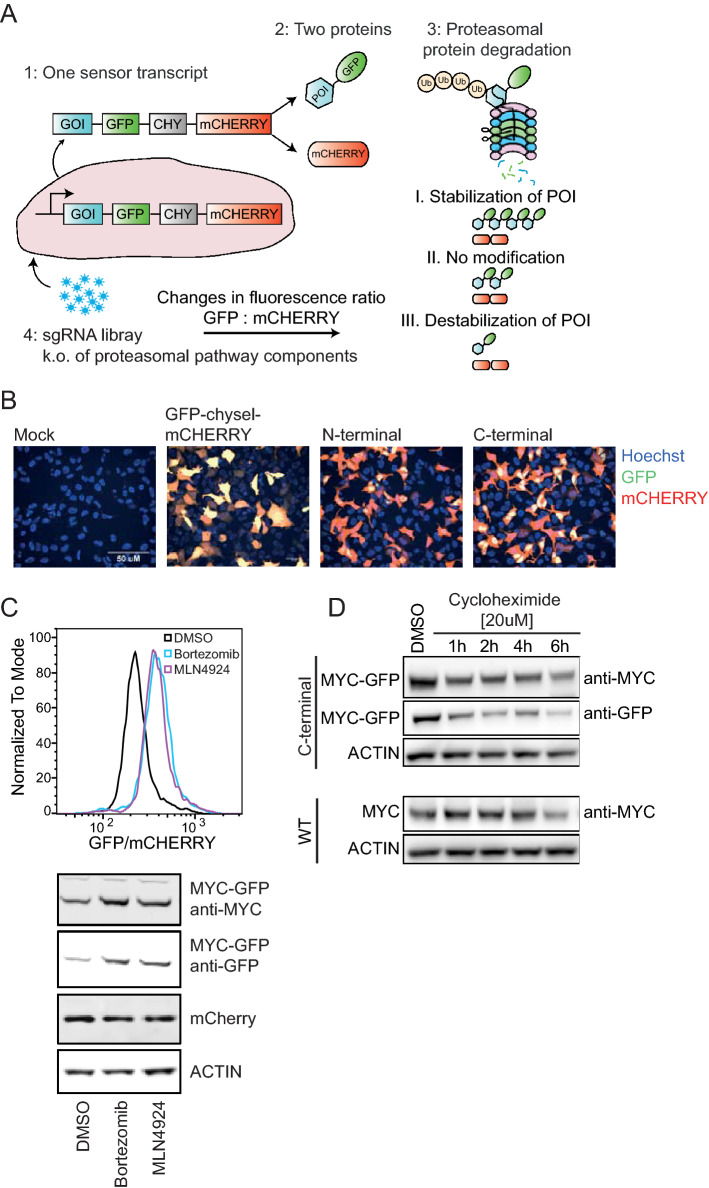
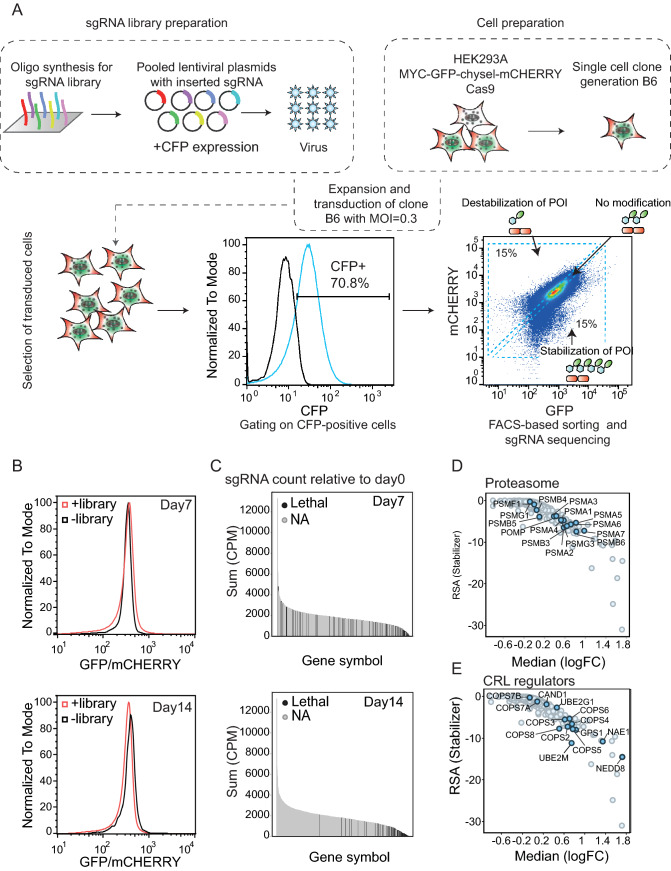
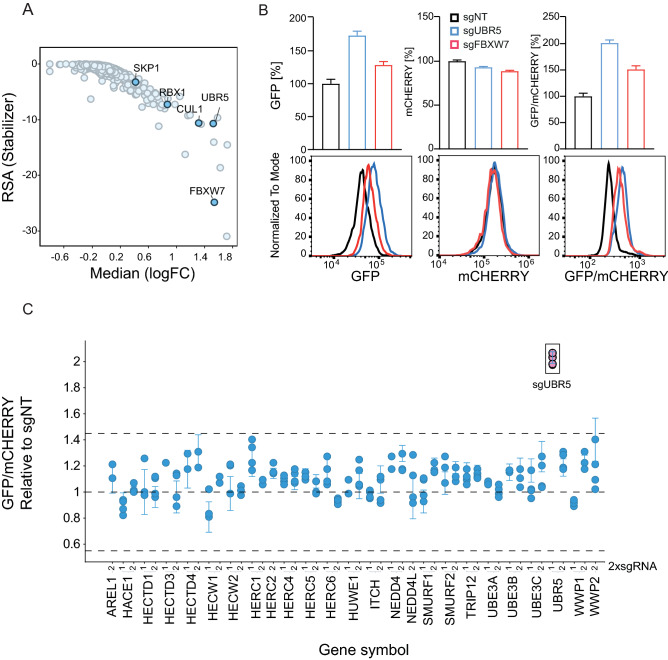
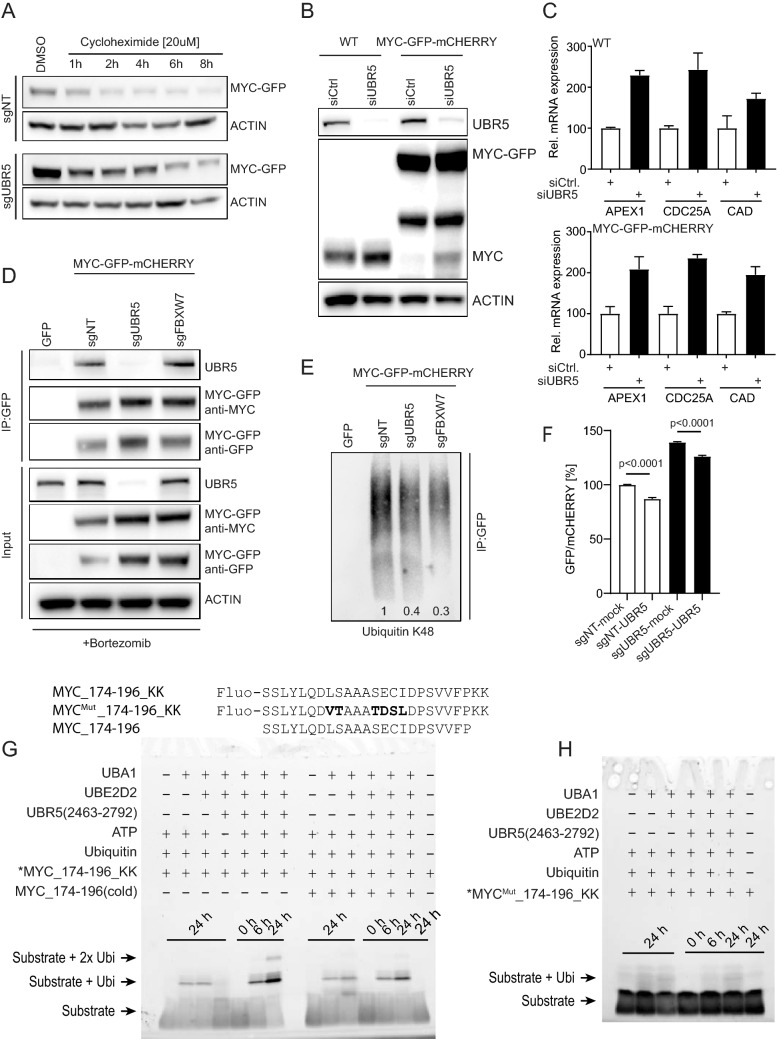
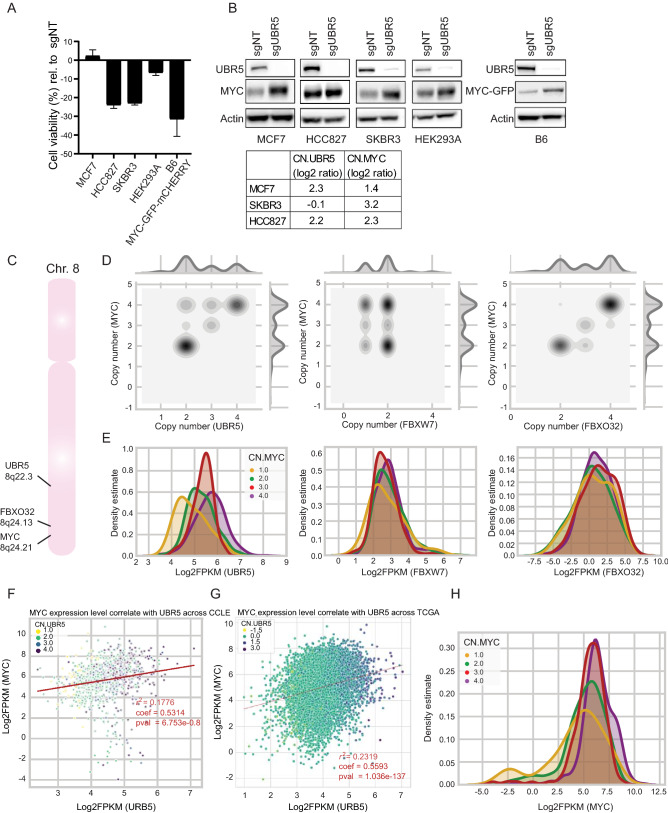
MYC oncoprotein is a multifunctional transcription factor that regulates the expression of a large number of genes involved in cellular growth, proliferation and metabolism. Altered MYC protein level lead to cellular transformation and tumorigenesis. MYC is deregulated in > 50% of human cancers, rendering it an attractive drug target. However, direct inhibition of this class of proteins using conventional small molecules is challenging due to their intrinsically disordered state. To discover novel posttranslational regulators of MYC protein stability and turnover, we established a genetic screen in mammalian cells by combining a fluorescent protein-based MYC abundance sensor, CRISPR/Cas9-based gene knockouts and next-generation sequencing. Our screen identifies UBR5, an E3 ligase of the HECT-type family, as a novel regulator of MYC degradation. Even in the presence of the well-described and functional MYC ligase, FBXW7, UBR5 depletion leads to accumulation of MYC in cells. We demonstrate interaction of UBR5 with MYC and reduced K48-linked ubiquitination of MYC upon loss of UBR5 in cells. Interestingly, in cancer cell lines with amplified MYC expression, depletion of UBR5 resulted in reduced cell survival, as a consequence of MYC stabilization. Finally, we show that MYC and UBR5 are co-amplified in more than 40% of cancer cells and that MYC copy number amplification correlates with enhanced transcriptional output of UBR5. This suggests that UBR5 acts as a buffer in MYC amplified settings and protects these cells from apoptosis.
Conflict of interest statement
All authors are or were employees and shareholders of Novartis Pharma AG.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
