

BAP1 mutant uveal melanoma is stratified by metabolic phenotypes with distinct vulnerability to metabolic inhibitors
- PMID: 33208912
- PMCID: PMC7856044
- DOI: 10.1038/s41388-020-01554-y
BAP1 mutant uveal melanoma is stratified by metabolic phenotypes with distinct vulnerability to metabolic inhibitors
Erratum in
-
Correction to: BAP1 mutant uveal melanoma is stratified by metabolic phenotypes with distinct vulnerability to metabolic inhibitors.Oncogene. 2021 Mar;40(9):1753. doi: 10.1038/s41388-021-01645-4. Oncogene. 2021. PMID: 33510355 Free PMC article. No abstract available.
Abstract
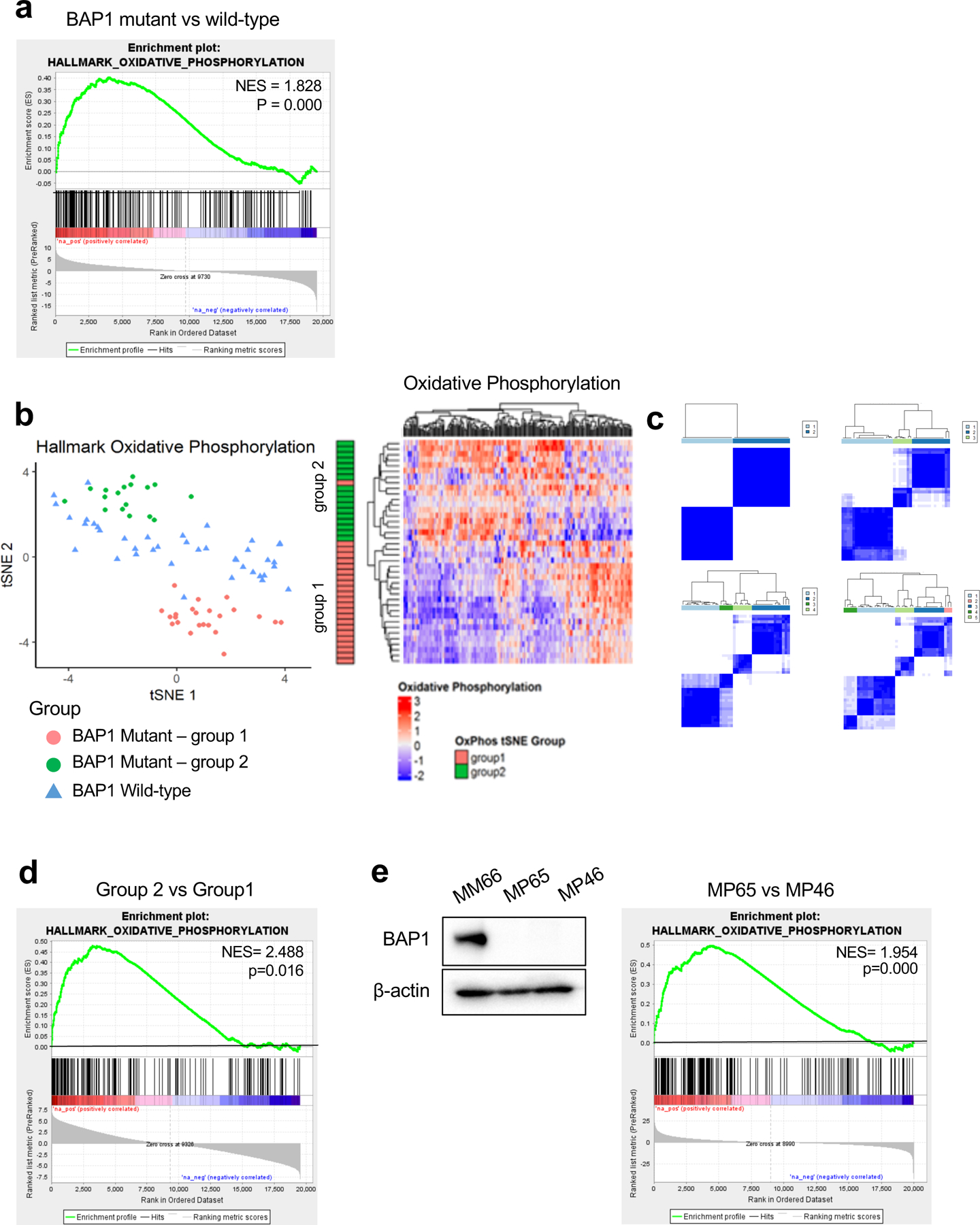
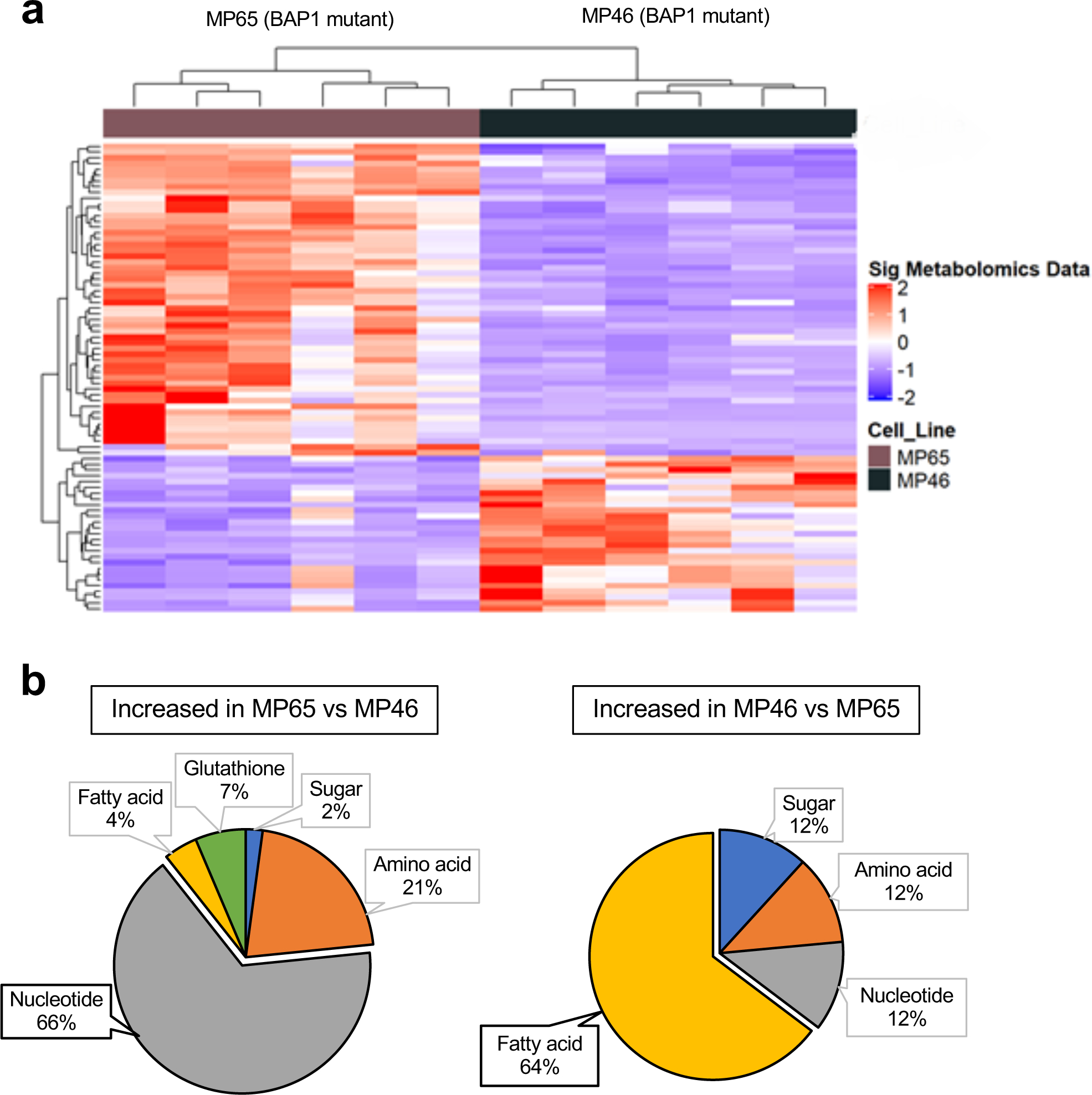
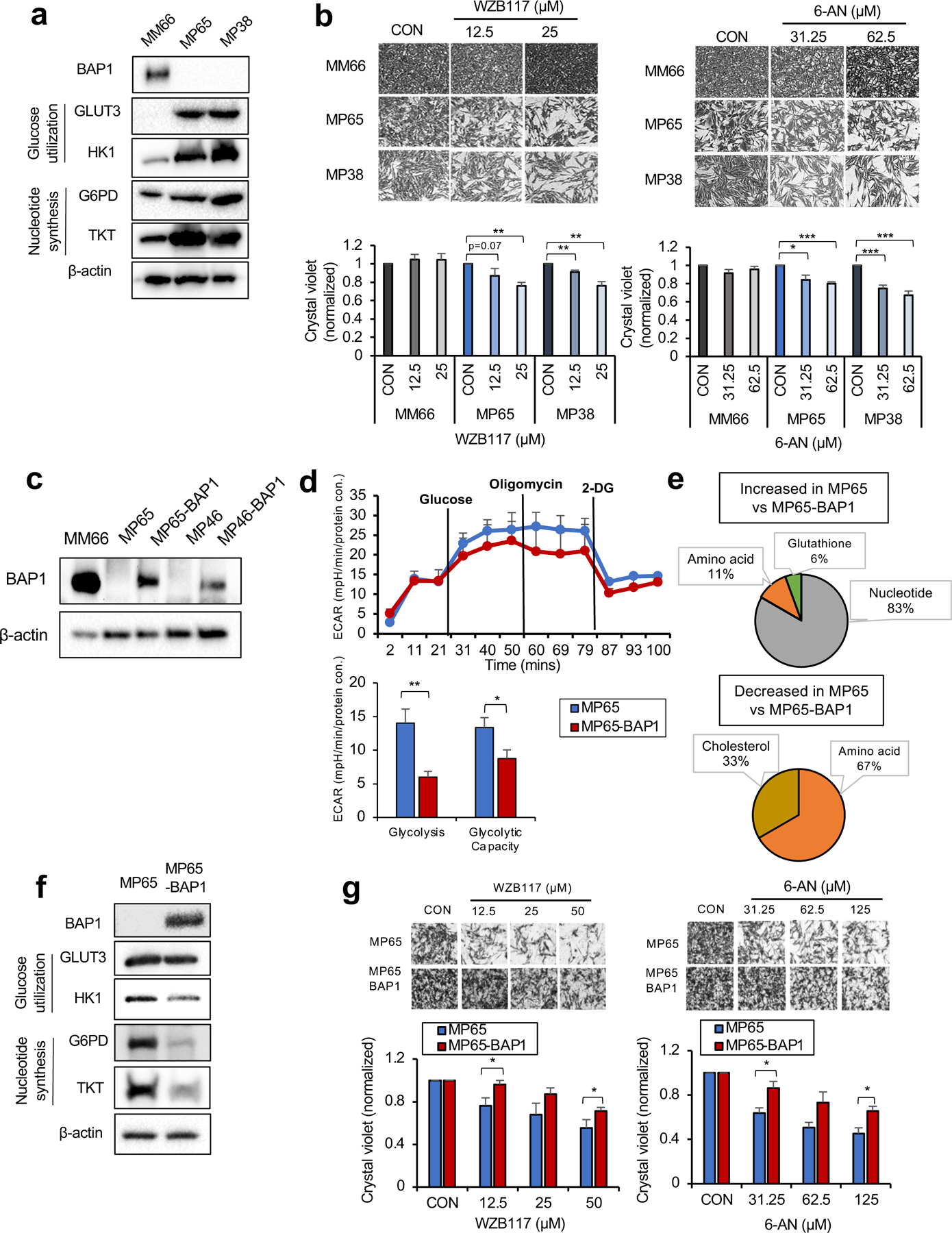
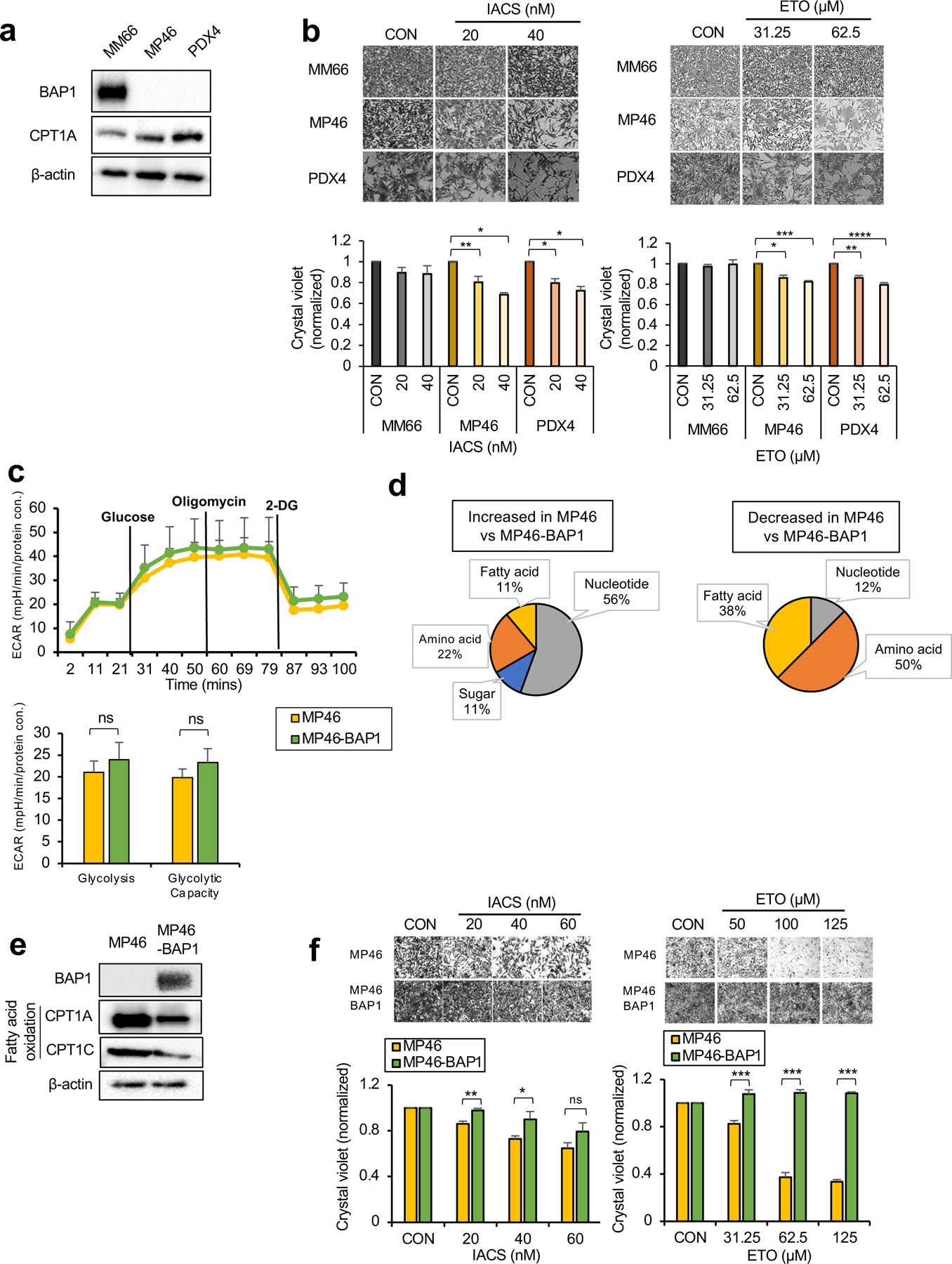
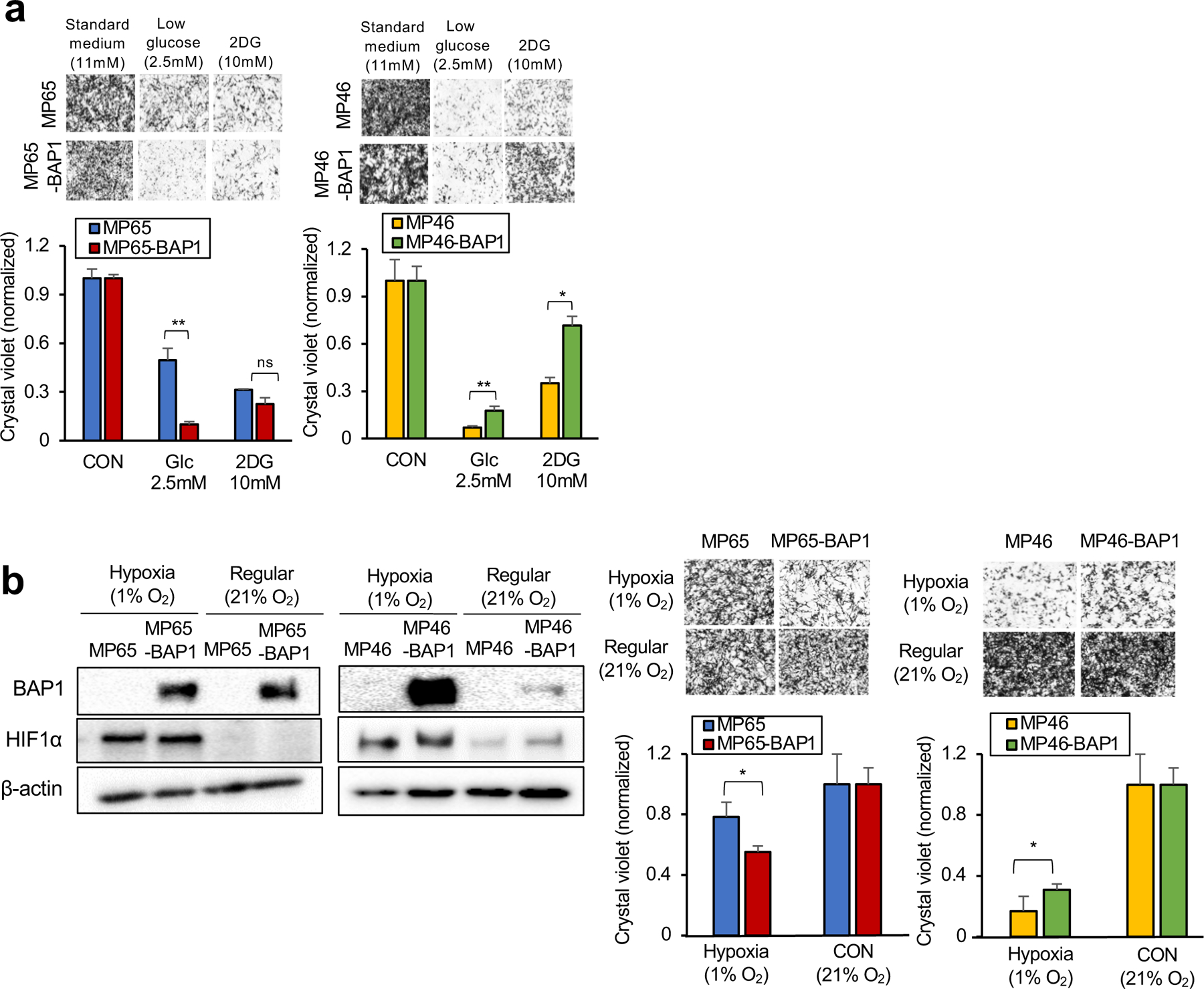
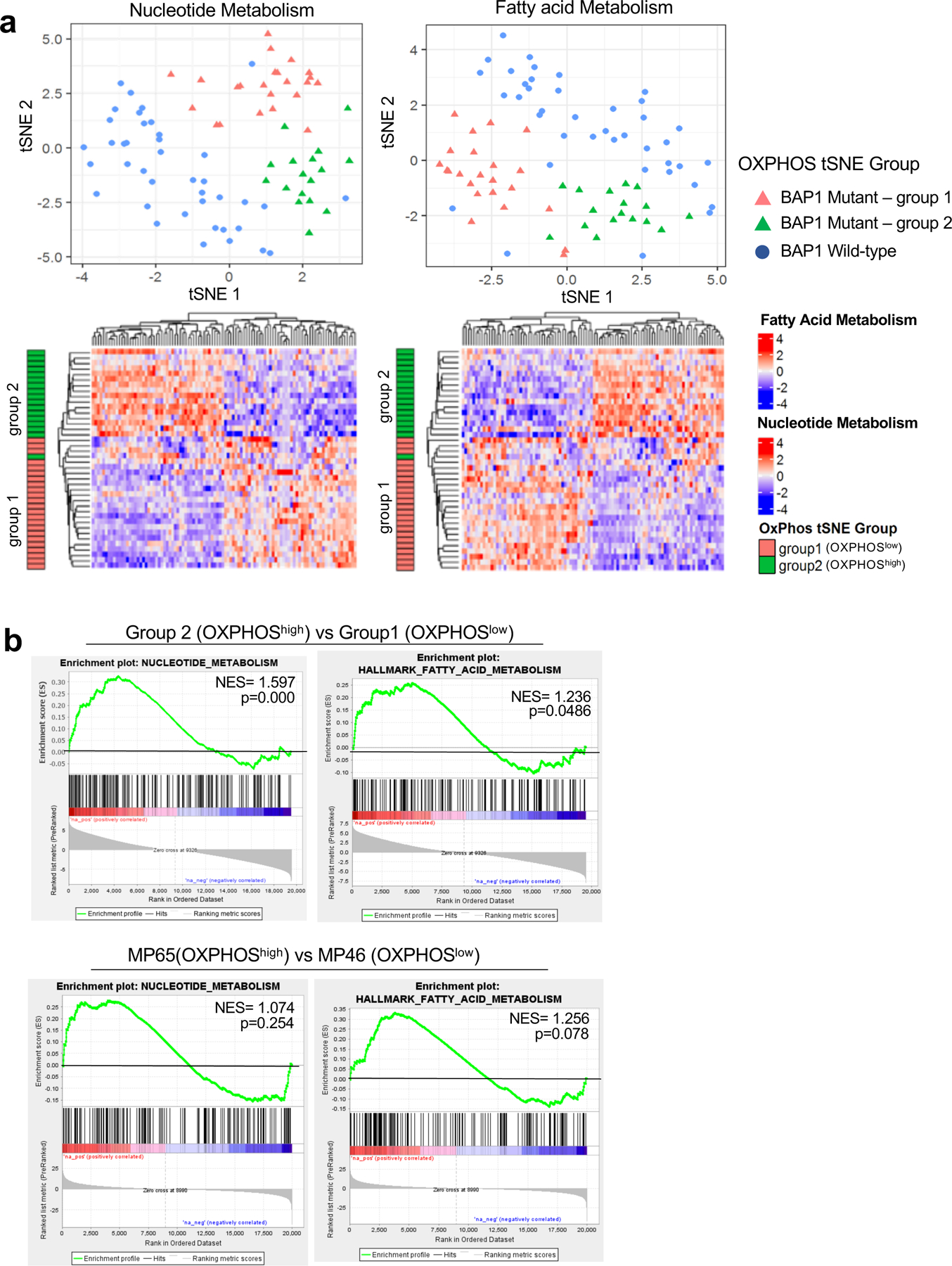
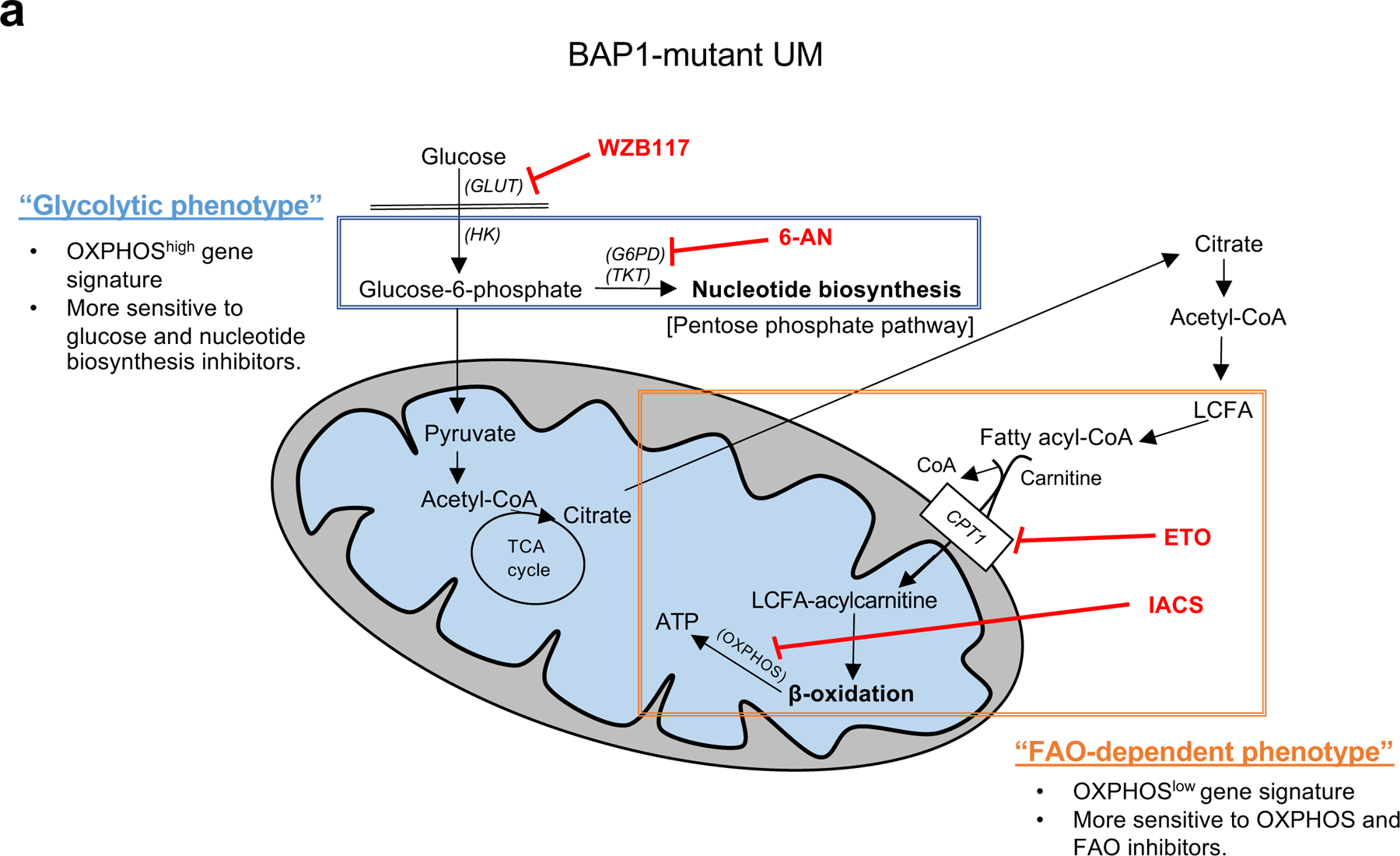
Cancer cell metabolism is a targetable vulnerability; however, a precise understanding of metabolic heterogeneity is required. Inactivating mutations in BRCA1-associated protein 1 (BAP1) are associated with metastasis in uveal melanoma (UM), the deadliest adult eye cancer. BAP1 functions in UM remain unclear. UM patient sample analysis divided BAP1 mutant UM tumors into two subgroups based on oxidative phosphorylation (OXPHOS) gene expression suggesting metabolic heterogeneity. Consistent with patient data, transcriptomic analysis of BAP1 mutant UM cell lines also showed OXPHOShigh or OXPHOSlow subgroups. Integrated RNA sequencing, metabolomics, and molecular analyses showed that OXPHOShigh BAP1 mutant UM cells utilize glycolytic and nucleotide biosynthesis pathways, whereas OXPHOSlow BAP1 mutant UM cells employ fatty acid oxidation. Furthermore, the two subgroups responded to different classes of metabolic suppressors. Our findings indicate that targeting cancer metabolism is a promising therapeutic option for BAP1 mutant UM; however, tailored approaches may be required due to metabolic heterogeneities.
Conflict of interest statement
Declaration of Interests:
A.E. Aplin reports receiving a commercial research grant from Pfizer Inc. (2013–2017) and has ownership interest in patent number 9880150. No potential conflicts of interest are disclosed by the other authors. J.W. Harbour is the inventor of intellectual property related to prognostic testing for uveal melanoma. He is a paid consultant for Castle Biosciences, licensee of this intellectual property, and he receives royalties from its commercialization.
Figures
References
-
- Jones NP, Schulze A. Targeting cancer metabolism–aiming at a tumour’s sweet-spot. Drug Discov Today. 2012;17(5–6):232–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
