

Loss of Adenylyl Cyclase 6 in Leptin Receptor-Expressing Stromal Cells Attenuates Loading-Induced Endosteal Bone Formation
- PMID: 33210061
- PMCID: PMC7657397
- DOI: 10.1002/jbm4.10408
Loss of Adenylyl Cyclase 6 in Leptin Receptor-Expressing Stromal Cells Attenuates Loading-Induced Endosteal Bone Formation
Abstract
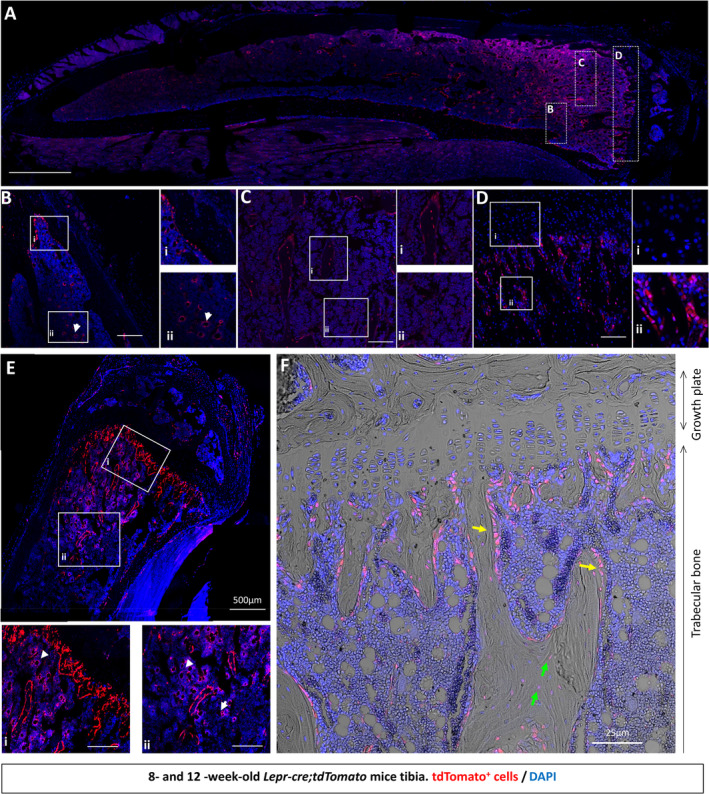
Bone marrow stromal/stem cells represent a quiescent cell population that replenish the osteoblast bone-forming cell pool with age and in response to injury, maintaining bone mass and repair. A potent mediator of stromal/stem cell differentiation in vitro and bone formation in vivo is physical loading, yet it still remains unclear whether loading-induced bone formation requires the osteogenic differentiation of these resident stromal/stem cells. Therefore, in this study, we utilized the leptin receptor (LepR) to identify and trace the contribution of bone marrow stromal cells to mechanoadaptation of bone in vivo. Twelve-week-old Lepr-cre;tdTomato mice were subjected to compressive tibia loading with an 11 N peak load for 40 cycles, every other day for 2 weeks. Histological analysis revealed that Lepr-cre;tdTomato+ cells arise perinatally around blood vessels and populate bone surfaces as lining cells or osteoblasts before a percentage undergo osteocytogenesis. Lepr-cre;tdTomato+ stromal cells within the marrow increase in abundance with age, but not following the application of tibial compressive loading. Mechanical loading induces an increase in bone mass and bone formation parameters, yet does not evoke an increase in Lepr-cre;tdTomato+ osteoblasts or osteocytes. To investigate whether adenylyl cyclase-6 (AC6) in LepR cells contributes to this mechanoadaptive response, Lepr-cre;tdTomato mice were further crossed with AC6 fl/fl mice to generate a LepR+ cell-specific knockout of AC6. These Lepr-cre;tdTomato;AC6 fl/fl animals have an attenuated response to compressive tibia loading, characterized by a deficient load-induced osteogenic response on the endosteal bone surface. This, therefore, shows that Lepr-cre;tdTomato+ cells contribute to short-term bone mechanoadaptation. © 2020 The Authors. JBMR Plus published by Wiley Periodicals LLC on behalf of American Society for Bone and Mineral Research.
Keywords: ADENYLYL CYCLASE 6; BONE ADAPTATION; IN VIVO MECHANICAL LOADING; MECHANOBIOLOGY; STEM CELLS.
© 2020 The Authors. JBMR Plus published by Wiley Periodicals LLC on behalf of American Society for Bone and Mineral Research.
Figures






Similar articles
-
From Marrow to Bone and Fat: Exploring the Multifaceted Roles of Leptin Receptor Positive Bone Marrow Mesenchymal Stromal Cells.Cells. 2024 May 24;13(11):910. doi: 10.3390/cells13110910. Cells. 2024. PMID: 38891042 Free PMC article. Review.
-
Leptin Receptor Promotes Adipogenesis and Reduces Osteogenesis by Regulating Mesenchymal Stromal Cells in Adult Bone Marrow.Cell Stem Cell. 2016 Jun 2;18(6):782-796. doi: 10.1016/j.stem.2016.02.015. Epub 2016 Mar 24. Cell Stem Cell. 2016. PMID: 27053299
-
Parathyroid Hormone Shifts Cell Fate of a Leptin Receptor-Marked Stromal Population from Adipogenic to Osteoblastic Lineage.J Bone Miner Res. 2019 Oct;34(10):1952-1963. doi: 10.1002/jbmr.3811. Epub 2019 Aug 2. J Bone Miner Res. 2019. PMID: 31173642
-
Leptin receptor-expressing cells represent a distinct subpopulation of notochord-derived cells and are essential for disc homoeostasis.J Orthop Translat. 2019 Dec 24;21:91-99. doi: 10.1016/j.jot.2019.11.005. eCollection 2020 Mar. J Orthop Translat. 2019. PMID: 32110508 Free PMC article.
-
Differentiation of committed osteoblast progenitors by octacalcium phosphate compared to calcium-deficient hydroxyapatite in Lepr-cre/Tomato mouse tibia.Acta Biomater. 2022 Apr 1;142:332-344. doi: 10.1016/j.actbio.2022.02.016. Epub 2022 Feb 18. Acta Biomater. 2022. PMID: 35183778
Cited by
-
Mechanobiology and Primary Cilium in the Pathophysiology of Bone Marrow Myeloproliferative Diseases.Int J Mol Sci. 2024 Aug 14;25(16):8860. doi: 10.3390/ijms25168860. Int J Mol Sci. 2024. PMID: 39201546 Free PMC article. Review.
-
Skeletal adaptation to mechanical cues during homeostasis and repair: the niche, cells, and molecular signaling.Front Physiol. 2023 Oct 17;14:1233920. doi: 10.3389/fphys.2023.1233920. eCollection 2023. Front Physiol. 2023. PMID: 37916223 Free PMC article. Review.
-
From Marrow to Bone and Fat: Exploring the Multifaceted Roles of Leptin Receptor Positive Bone Marrow Mesenchymal Stromal Cells.Cells. 2024 May 24;13(11):910. doi: 10.3390/cells13110910. Cells. 2024. PMID: 38891042 Free PMC article. Review.
-
Osteocytes and Primary Cilia.Curr Osteoporos Rep. 2023 Dec;21(6):719-730. doi: 10.1007/s11914-023-00819-1. Epub 2023 Sep 8. Curr Osteoporos Rep. 2023. PMID: 37682373 Free PMC article. Review.
References
-
- Frost HM. Skeletal structural adaptations to mechanical usage (SATMU): 1. Redefining Wolff's law: the bone modeling problem. Anat Rec. 1990;226(4):403–13. - PubMed
-
- Stavenschi E, Labour MN, Hoey DA. Oscillatory fluid flow induces the osteogenic lineage commitment of mesenchymal stem cells: the effect of shear stress magnitude, frequency and duration. J Biomech. 2017;55:99–106. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous