

Evolution of multicellular life cycles under costly fragmentation
- PMID: 33211685
- PMCID: PMC7714367
- DOI: 10.1371/journal.pcbi.1008406
Evolution of multicellular life cycles under costly fragmentation
Abstract
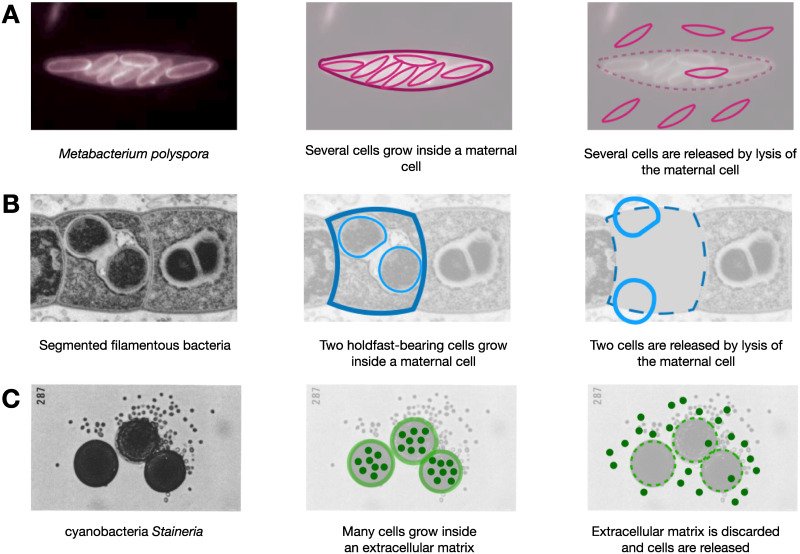
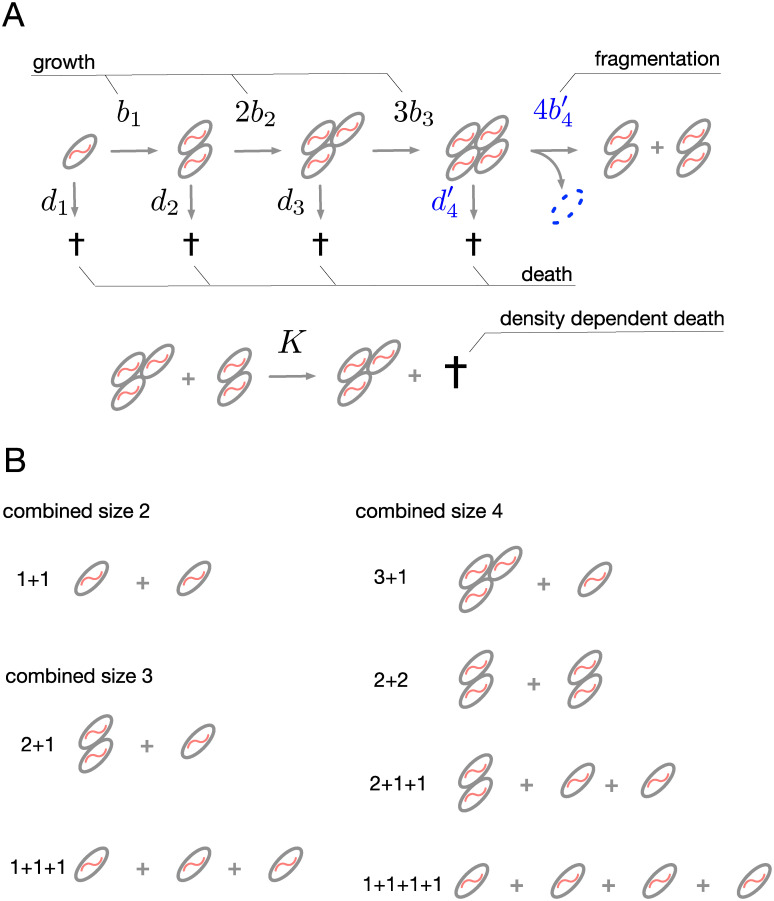
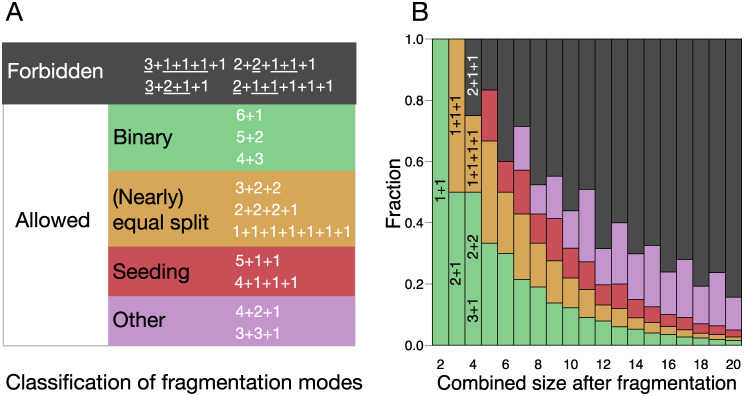
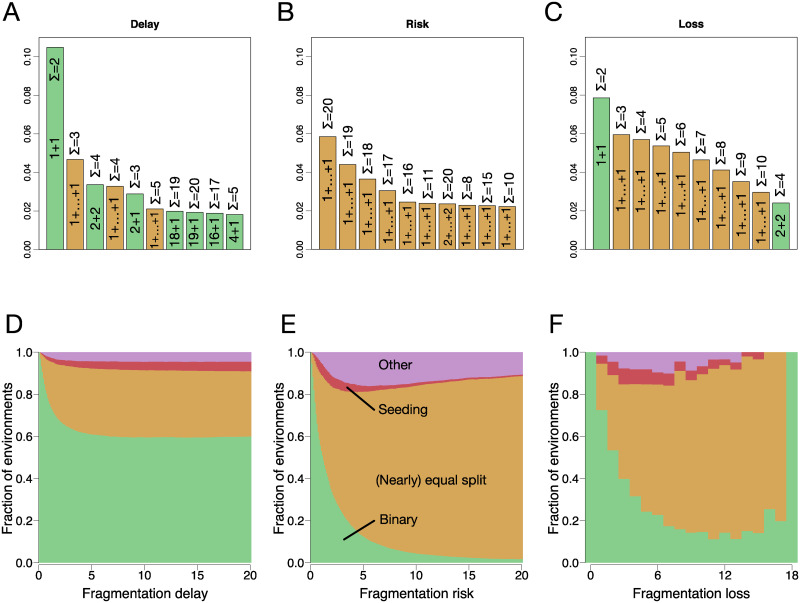
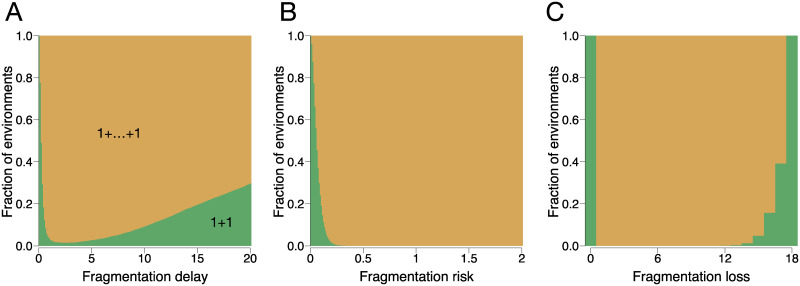
A fascinating wealth of life cycles is observed in biology, from unicellularity to the concerted fragmentation of multicellular units. However, the understanding of factors driving their evolution is still limited. We show that costs of fragmentation have a major impact on the evolution of life cycles due to their influence on the growth rates of the associated populations. We model a group structured population of undifferentiated cells, where cell clusters reproduce by fragmentation. Fragmentation events are associated with a cost expressed by either a fragmentation delay, an additional risk, or a cell loss. The introduction of such fragmentation costs vastly increases the set of possible life cycles. Based on these findings, we suggest that the evolution of life cycles involving splitting into multiple offspring can be directly associated with the fragmentation cost. Moreover, the impact of this cost alone is strong enough to drive the emergence of multicellular units that eventually split into many single cells, even under scenarios that strongly disfavour collectives compared to solitary individuals.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Fragmentation modes and the evolution of life cycles.PLoS Comput Biol. 2017 Nov 22;13(11):e1005860. doi: 10.1371/journal.pcbi.1005860. eCollection 2017 Nov. PLoS Comput Biol. 2017. PMID: 29166656 Free PMC article.
-
Interacting cells driving the evolution of multicellular life cycles.PLoS Comput Biol. 2019 May 14;15(5):e1006987. doi: 10.1371/journal.pcbi.1006987. eCollection 2019 May. PLoS Comput Biol. 2019. PMID: 31086369 Free PMC article.
-
Evolution of simple multicellular life cycles in dynamic environments.J R Soc Interface. 2019 May 31;16(154):20190054. doi: 10.1098/rsif.2019.0054. J R Soc Interface. 2019. PMID: 31088261 Free PMC article.
-
Trematode life cycles: short is sweet?Trends Parasitol. 2002 Apr;18(4):176-83. doi: 10.1016/s1471-4922(02)02262-6. Trends Parasitol. 2002. PMID: 11998706 Review.
-
The evolution of growth trajectories: what limits growth rate?Biol Rev Camb Philos Soc. 2011 Feb;86(1):97-116. doi: 10.1111/j.1469-185X.2010.00136.x. Biol Rev Camb Philos Soc. 2011. PMID: 20394607 Review.
Cited by
-
Evolution of reproductive strategies in incipient multicellularity.J R Soc Interface. 2022 Mar;19(188):20210716. doi: 10.1098/rsif.2021.0716. Epub 2022 Mar 2. J R Soc Interface. 2022. PMID: 35232276 Free PMC article.
-
Eco-evolutionary dynamics of clonal multicellular life cycles.Elife. 2022 Sep 13;11:e78822. doi: 10.7554/eLife.78822. Elife. 2022. PMID: 36099169 Free PMC article.
-
Multilevel selection favors fragmentation modes that maintain cooperative interactions in multispecies communities.PLoS Comput Biol. 2021 Sep 13;17(9):e1008896. doi: 10.1371/journal.pcbi.1008896. eCollection 2021 Sep. PLoS Comput Biol. 2021. PMID: 34516543 Free PMC article.
-
Cell size and selection for stress-induced cell fusion in unicellular eukaryotes.PLoS Comput Biol. 2025 Apr 8;21(4):e1012418. doi: 10.1371/journal.pcbi.1012418. eCollection 2025. PLoS Comput Biol. 2025. PMID: 40198726 Free PMC article.
-
Alternating selection for dispersal and multicellularity favors regulated life cycles.Curr Biol. 2023 May 8;33(9):1809-1817.e3. doi: 10.1016/j.cub.2023.03.031. Epub 2023 Apr 4. Curr Biol. 2023. PMID: 37019107 Free PMC article.
References
-
- Stearns SC. The evolution of life histories. Oxford University Press, NY; 1992.
MeSH terms
Supplementary concepts
LinkOut - more resources
Full Text Sources
