

How directed evolution reshapes the energy landscape in an enzyme to boost catalysis
- PMID: 33214289
- PMCID: PMC9616100
- DOI: 10.1126/science.abd3623
How directed evolution reshapes the energy landscape in an enzyme to boost catalysis
Abstract
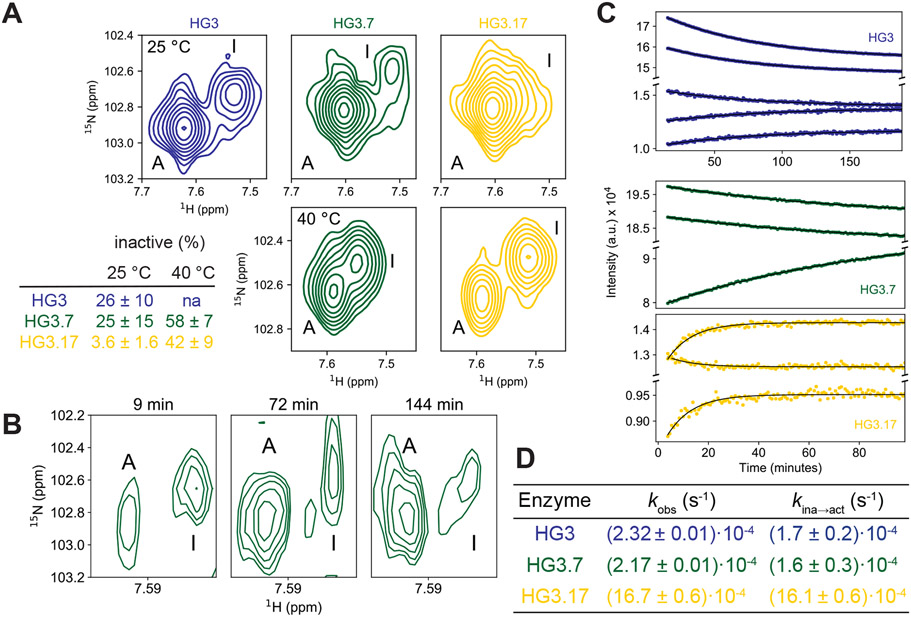
The advent of biocatalysts designed computationally and optimized by laboratory evolution provides an opportunity to explore molecular strategies for augmenting catalytic function. Applying a suite of nuclear magnetic resonance, crystallography, and stopped-flow techniques to an enzyme designed for an elementary proton transfer reaction, we show how directed evolution gradually altered the conformational ensemble of the protein scaffold to populate a narrow, highly active conformational ensemble and accelerate this transformation by nearly nine orders of magnitude. Mutations acquired during optimization enabled global conformational changes, including high-energy backbone rearrangements, that cooperatively organized the catalytic base and oxyanion stabilizer, thus perfecting transition-state stabilization. The development of protein catalysts for many chemical transformations could be facilitated by explicitly sampling conformational substates during design and specifically stabilizing productive substates over all unproductive conformations.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures



References
-
- Kiss G, Celebi-Olcum N, Moretti R, Baker D, Houk KN, Computational enzyme design. Angew Chem Int Ed Engl 52, 5700–5725 (2013). - PubMed
-
- Kries H, Blomberg R, Hilvert D, De novo enzymes by computational design. Curr Opin Chem Biol 17, 221–228 (2013). - PubMed
-
- Blomberg R et al., Precision is essential for efficient catalysis in an evolved Kemp eliminase. Nature 503, 418–421 (2013). - PubMed
-
- Obexer R et al., Emergence of a catalytic tetrad during evolution of a highly active artificial aldolase. Nat Chem 9, 50–56 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
