

Protein Conformational Dynamics upon Association with the Surfaces of Lipid Membranes and Engineered Nanoparticles: Insights from Electron Paramagnetic Resonance Spectroscopy
- PMID: 33218036
- PMCID: PMC7698768
- DOI: 10.3390/molecules25225393
Protein Conformational Dynamics upon Association with the Surfaces of Lipid Membranes and Engineered Nanoparticles: Insights from Electron Paramagnetic Resonance Spectroscopy
Abstract
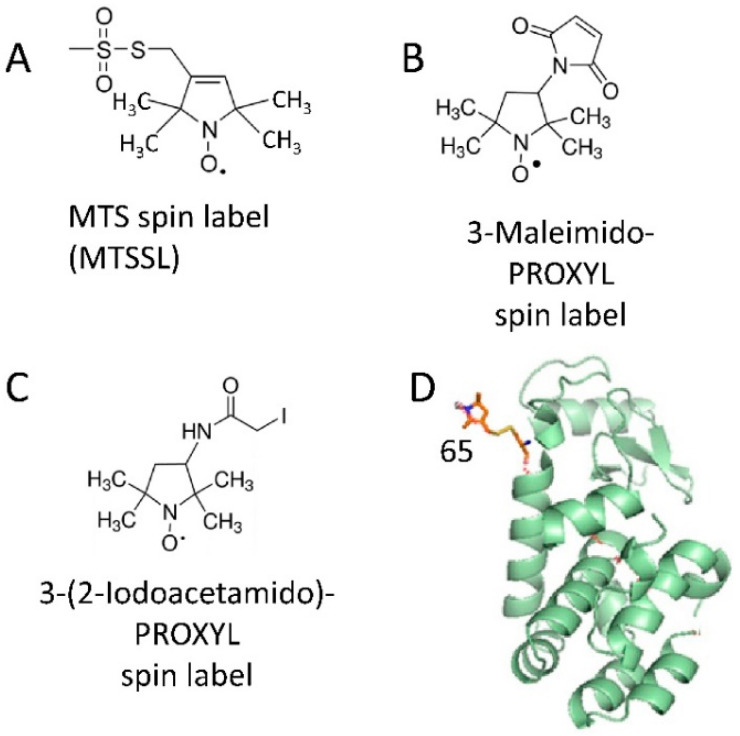
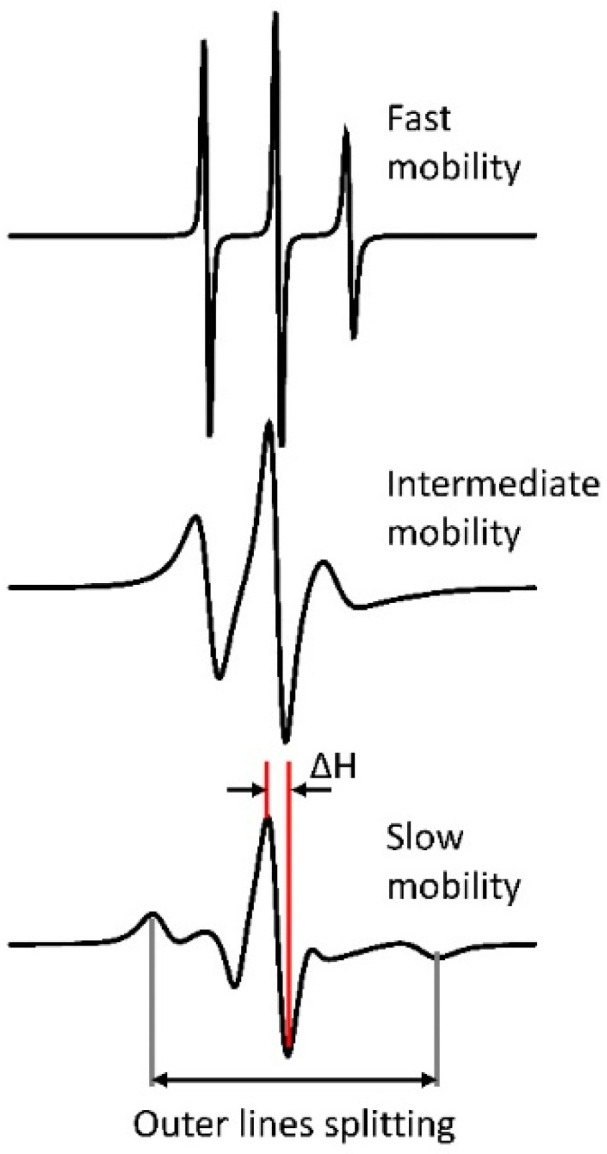
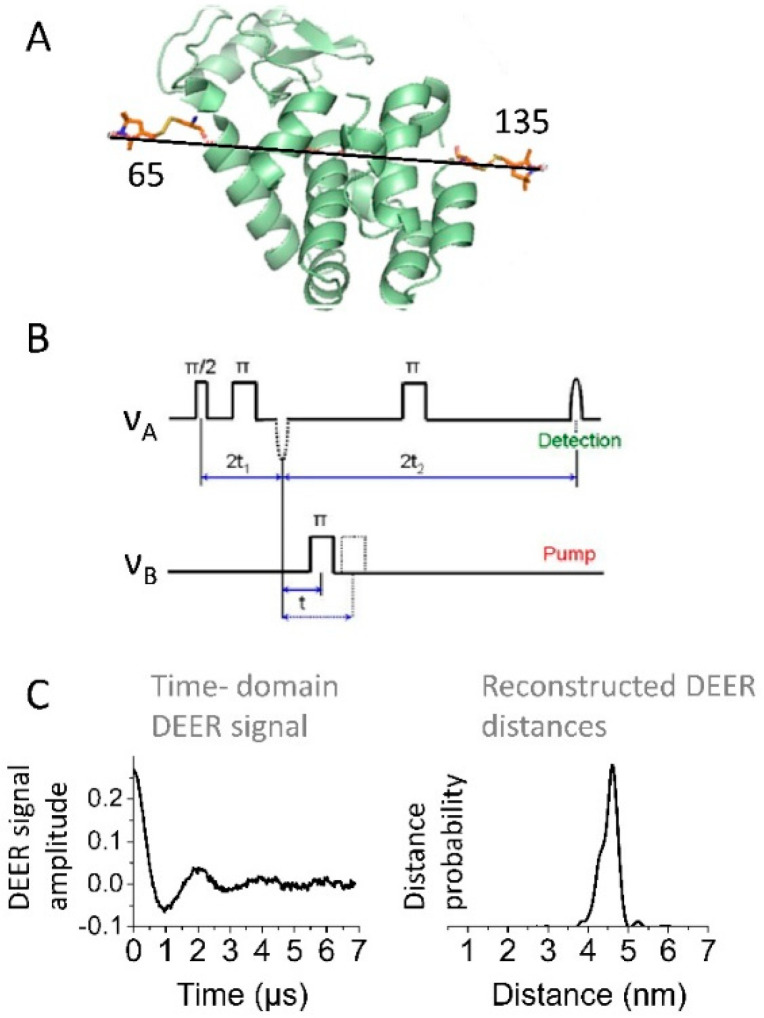
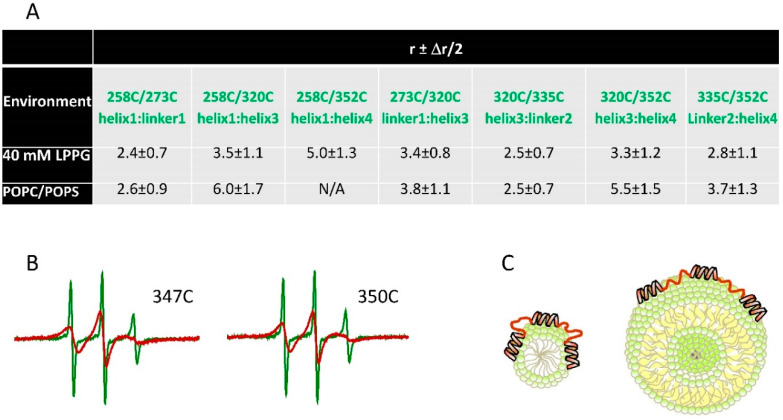
Detailed study of conformational rearrangements and dynamics of proteins is central to our understanding of their physiological functions and the loss of function. This review outlines the applications of the electron paramagnetic resonance (EPR) technique to study the structural aspects of proteins transitioning from a solution environment to the states in which they are associated with the surfaces of biological membranes or engineered nanoobjects. In the former case these structural transitions generally underlie functional protein states. The latter case is mostly relevant to the application of protein immobilization in biotechnological industries, developing methods for protein purification, etc. Therefore, evaluating the stability of the protein functional state is particularly important. EPR spectroscopy in the form of continuous-wave EPR or pulse EPR distance measurements in conjunction with protein spin labeling provides highly versatile and sensitive tools to characterize the changes in protein local dynamics as well as large conformational rearrangements. The technique can be widely utilized in studies of both protein-membrane and engineered nanoobject-protein complexes.
Keywords: EPR spectroscopy; lipid membranes; protein conformation; protein-surface association; surface-immobilized protein.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Structural Flexibility of Tau in Its Interaction with Microtubules as Viewed by Site-Directed Spin Labeling EPR Spectroscopy.Methods Mol Biol. 2024;2754:55-75. doi: 10.1007/978-1-0716-3629-9_3. Methods Mol Biol. 2024. PMID: 38512660
-
EPR Techniques to Probe Insertion and Conformation of Spin-Labeled Proteins in Lipid Bilayers.Methods Mol Biol. 2019;2003:493-528. doi: 10.1007/978-1-4939-9512-7_21. Methods Mol Biol. 2019. PMID: 31218631
-
Studying Lipid-Protein Interactions with Electron Paramagnetic Resonance Spectroscopy of Spin-Labeled Lipids.Methods Mol Biol. 2019;2003:529-561. doi: 10.1007/978-1-4939-9512-7_22. Methods Mol Biol. 2019. PMID: 31218632
-
Nitroxide spin labels and EPR spectroscopy: A powerful association for protein dynamics studies.Biochim Biophys Acta Proteins Proteom. 2021 Jul;1869(7):140653. doi: 10.1016/j.bbapap.2021.140653. Epub 2021 Mar 20. Biochim Biophys Acta Proteins Proteom. 2021. PMID: 33757896 Review.
-
Elucidating the design principles of photosynthetic electron-transfer proteins by site-directed spin labeling EPR spectroscopy.Biochim Biophys Acta. 2016 May;1857(5):548-556. doi: 10.1016/j.bbabio.2015.08.009. Epub 2015 Sep 1. Biochim Biophys Acta. 2016. PMID: 26334844 Review.
Cited by
-
Bone Formation on Murine Cranial Bone by Injectable Cross-Linked Hyaluronic Acid Containing Nano-Hydroxyapatite and Bone Morphogenetic Protein.Polymers (Basel). 2022 Dec 8;14(24):5368. doi: 10.3390/polym14245368. Polymers (Basel). 2022. PMID: 36559734 Free PMC article.
-
Protein Adsorption and Conformational Changes.Molecules. 2021 Nov 23;26(23):7079. doi: 10.3390/molecules26237079. Molecules. 2021. PMID: 34885661 Free PMC article.
-
Engineered Chimera Protein Constructs to Facilitate the Production of Heterologous Transmembrane Proteins in E. coli.Int J Mol Sci. 2024 Feb 16;25(4):2354. doi: 10.3390/ijms25042354. Int J Mol Sci. 2024. PMID: 38397029 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
