

Acid Sphingomyelinase Impacts Canonical Transient Receptor Potential Channels 6 (TRPC6) Activity in Primary Neuronal Systems
- PMID: 33218173
- PMCID: PMC7698877
- DOI: 10.3390/cells9112502
Acid Sphingomyelinase Impacts Canonical Transient Receptor Potential Channels 6 (TRPC6) Activity in Primary Neuronal Systems
Abstract
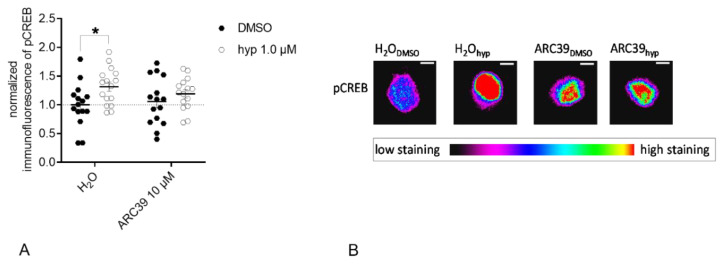
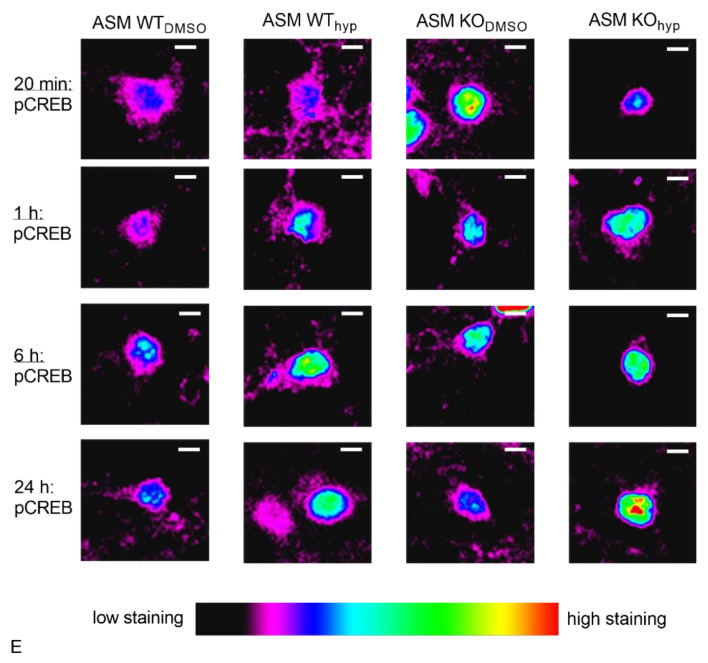
: The acid sphingomyelinase (ASM)/ceramide system exhibits a crucial role in the pathology of major depressive disorder (MDD). ASM hydrolyzes the abundant membrane lipid sphingomyelin to ceramide that regulates the clustering of membrane proteins via microdomain and lipid raft organization. Several commonly used antidepressants, such as fluoxetine, rely on the functional inhibition of ASM in terms of their antidepressive pharmacological effects. Transient receptor potential canonical 6 (TRPC6) ion channels are located in the plasma membrane of neurons and serve as receptors for hyperforin, a phytochemical constituent of the antidepressive herbal remedy St. John's wort. TRPC6 channels are involved in the regulation of neuronal plasticity, which likely contributes to their antidepressant effect. In this work, we investigated the impact of reduced ASM activity on the TRPC6 function in neurons. A lipidomic analysis of cortical brain tissue of ASM deficient mice revealed a decrease in ceramide/sphingomyelin molar ratio and an increase in sphingosine. In neurons with ASM deletion, hyperforin-mediated Ca2+-influx via TRPC6 was decreased. Consequently, downstream activation of nuclear phospho-cAMP response element-binding protein (pCREB) was changed, a transcriptional factor involved in neuronal plasticity. Our study underlines the importance of balanced ASM activity, as well as sphingolipidome composition for optimal TRPC6 function. A better understanding of the interaction of the ASM/ceramide and TRPC6 systems could help to draw conclusions about the pathology of MDD.
Keywords: acid sphingomyelinase; major depressive disorder; sphingolipids; trpc6.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures







References
-
- Schneider P.B., Kennedy E.P. Sphingomyelinase in normal human spleens and in spleens from subjects with Niemann-Pick disease. J. Lipid Res. 1967;8:202–209. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
