

CMT-3 targets different α-synuclein aggregates mitigating their toxic and inflammogenic effects
- PMID: 33219264
- PMCID: PMC7679368
- DOI: 10.1038/s41598-020-76927-0
CMT-3 targets different α-synuclein aggregates mitigating their toxic and inflammogenic effects
Abstract
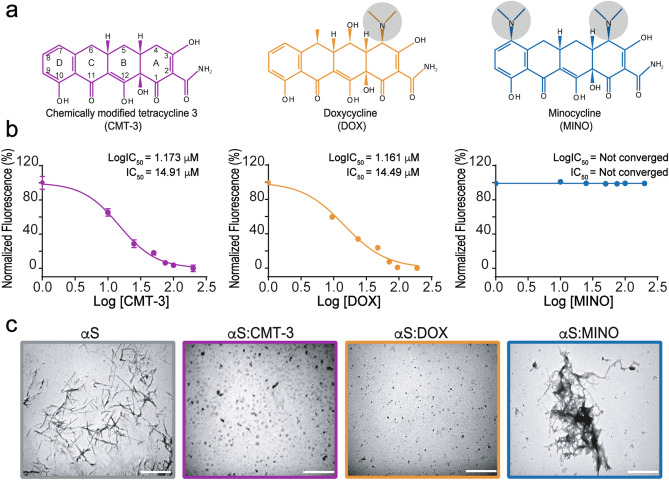
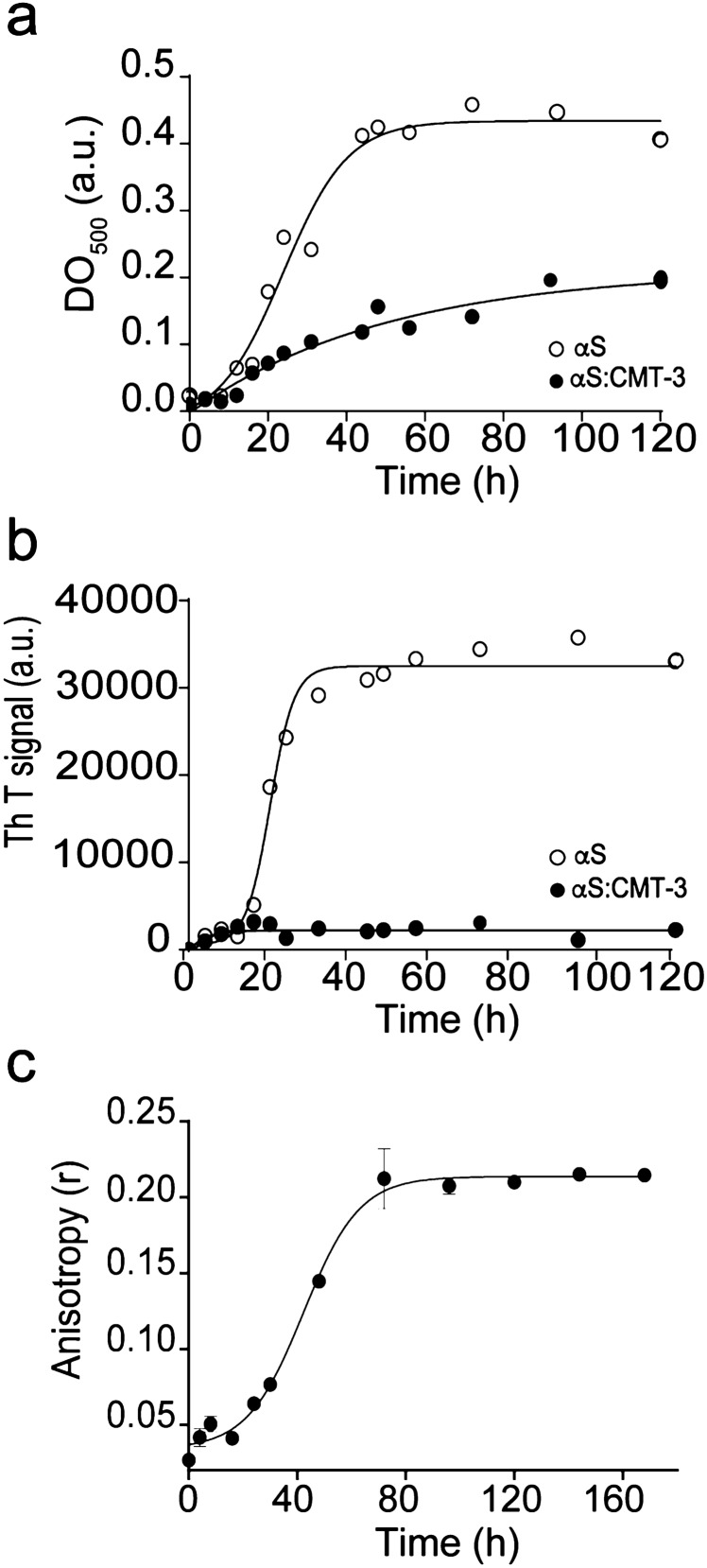
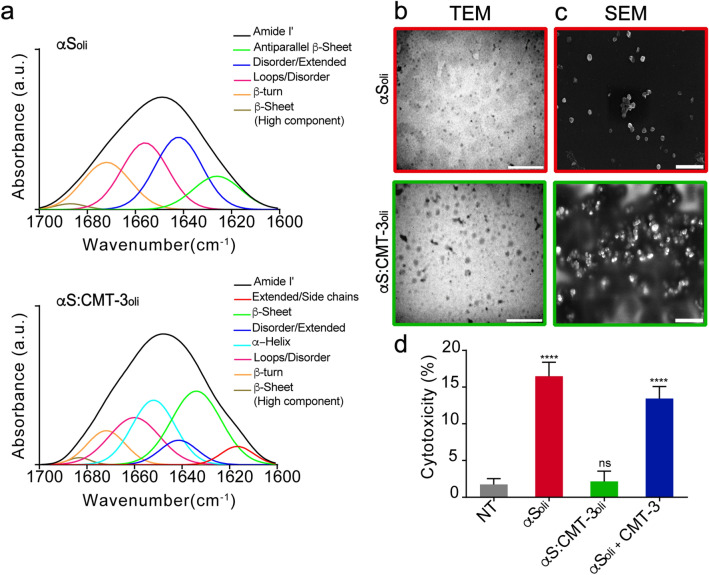
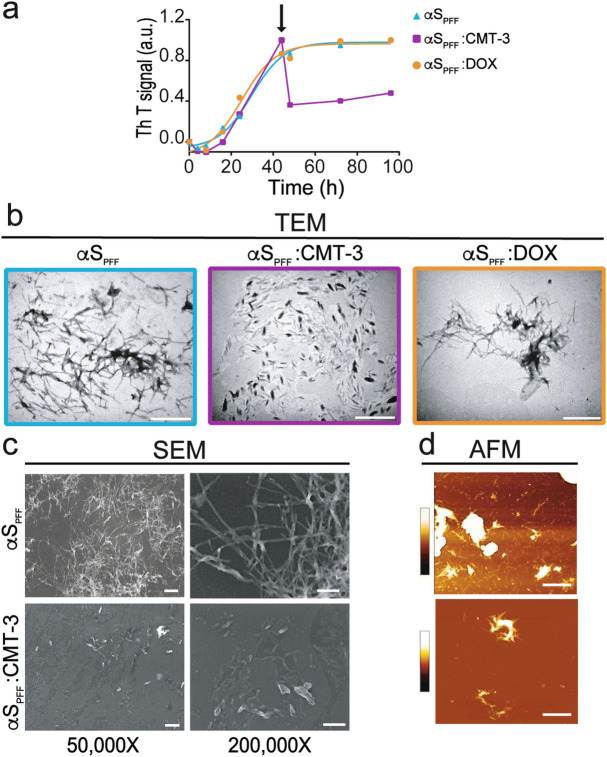
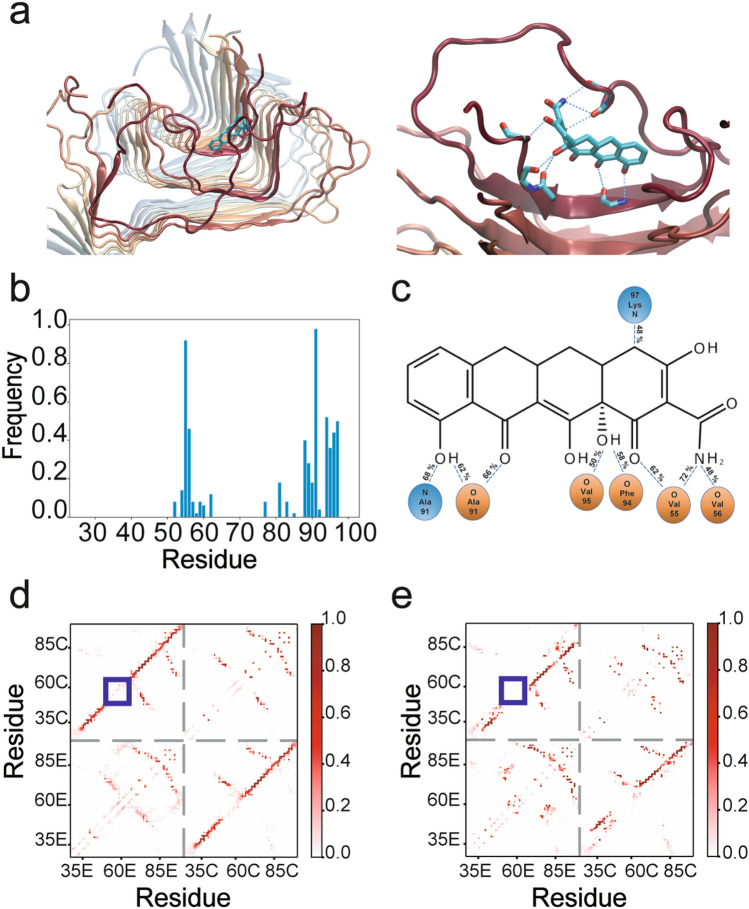
Parkinson's disease (PD) is a neurodegenerative disorder for which only symptomatic treatments are available. Repurposing drugs that target α-synuclein aggregation, considered one of the main drivers of PD progression, could accelerate the development of disease-modifying therapies. In this work, we focused on chemically modified tetracycline 3 (CMT-3), a derivative with reduced antibiotic activity that crosses the blood-brain barrier and is pharmacologically safe. We found that CMT-3 inhibited α-synuclein amyloid aggregation and led to the formation of non-toxic molecular species, unlike minocycline. Furthermore, CMT-3 disassembled preformed α-synuclein amyloid fibrils into smaller fragments that were unable to seed in subsequent aggregation reactions. Most interestingly, disaggregated species were non-toxic and less inflammogenic on brain microglial cells. Finally, we modelled the interactions between CMT-3 and α-synuclein aggregates by molecular simulations. In this way, we propose a mechanism for fibril disassembly. Our results place CMT-3 as a potential disease modifier for PD and possibly other synucleinopathies.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
