

Munc13 binds and recruits SNAP25 to chaperone SNARE complex assembly
- PMID: 33222163
- PMCID: PMC8068094
- DOI: 10.1002/1873-3468.14006
Munc13 binds and recruits SNAP25 to chaperone SNARE complex assembly
Abstract
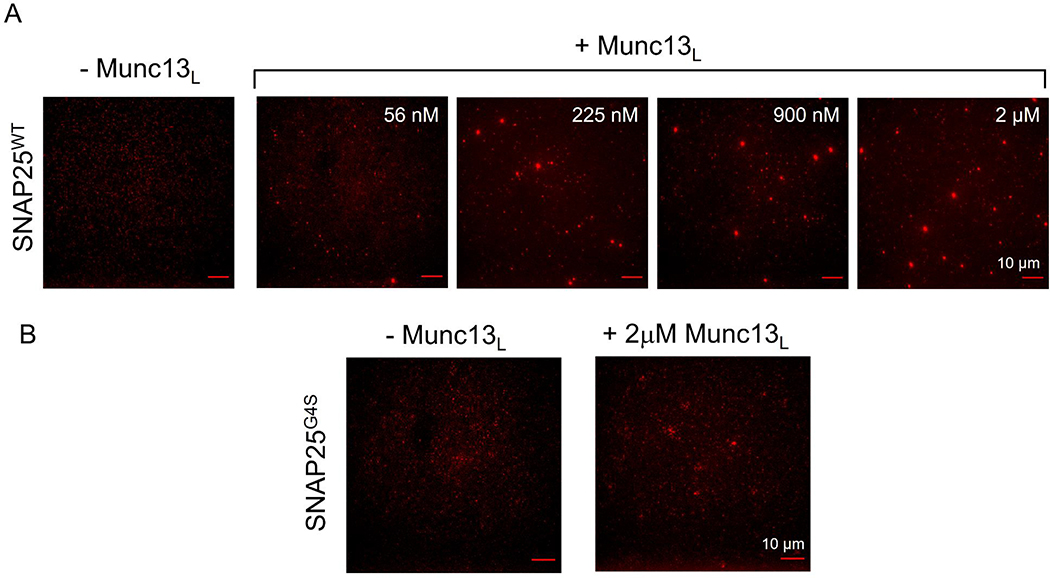
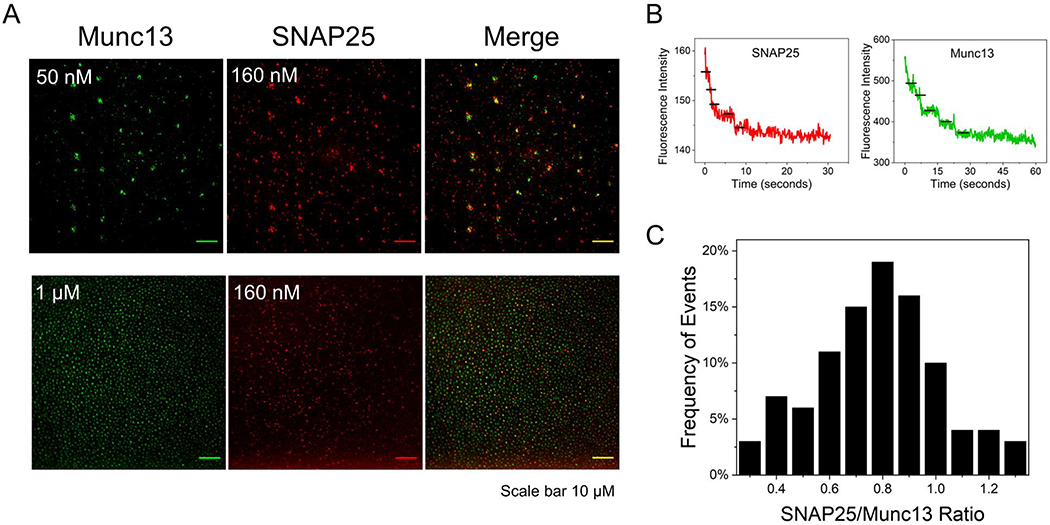
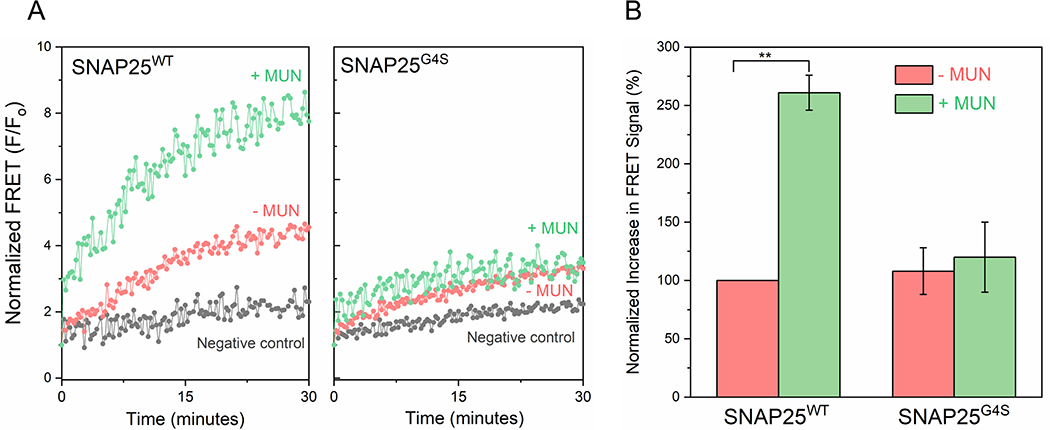
Synaptic vesicle fusion is mediated by SNARE proteins-VAMP2 on the vesicle and Syntaxin-1/SNAP25 on the presynaptic membrane. Chaperones Munc18-1 and Munc13-1 cooperatively catalyze SNARE assembly via an intermediate 'template' complex containing Syntaxin-1 and VAMP2. How SNAP25 enters this reaction remains a mystery. Here, we report that Munc13-1 recruits SNAP25 to initiate the ternary SNARE complex assembly by direct binding, as judged by bulk FRET spectroscopy and single-molecule optical tweezer studies. Detailed structure-function analyses show that the binding is mediated by the Munc13-1 MUN domain and is specific for the SNAP25 'linker' region that connects the two SNARE motifs. Consequently, freely diffusing SNAP25 molecules on phospholipid bilayers are concentrated and bound in ~ 1 : 1 stoichiometry by the self-assembled Munc13-1 nanoclusters.
© 2020 Federation of European Biochemical Societies.
Figures


References
-
- Rizo J and Xu J, The Synaptic Vesicle Release Machinery. Annu Rev Biophys, 2015. 44: p. 339–67. - PubMed
-
- Sollner T, et al. , A protein assembly-disassembly pathway in vitro that may correspond to sequential steps of synaptic vesicle docking, activation, and fusion. Cell, 1993. 75(3): p. 409–18. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
