

Mast cells and IgE in defense against lethality of venoms: Possible "benefit" of allergy[]
- PMID: 33224714
- PMCID: PMC7673288
- DOI: 10.1007/s40629-020-00118-6
Mast cells and IgE in defense against lethality of venoms: Possible "benefit" of allergy[]
Abstract
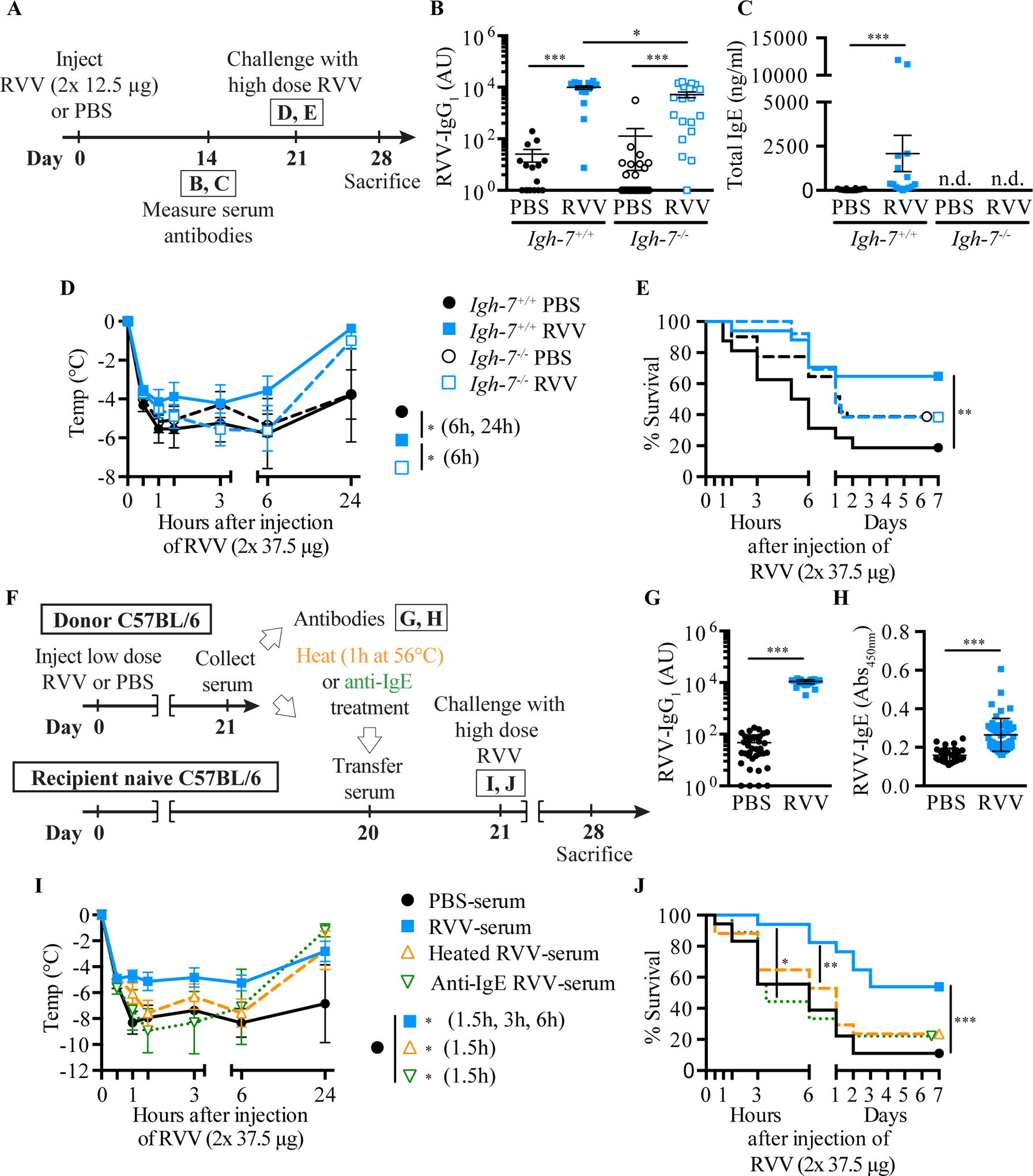
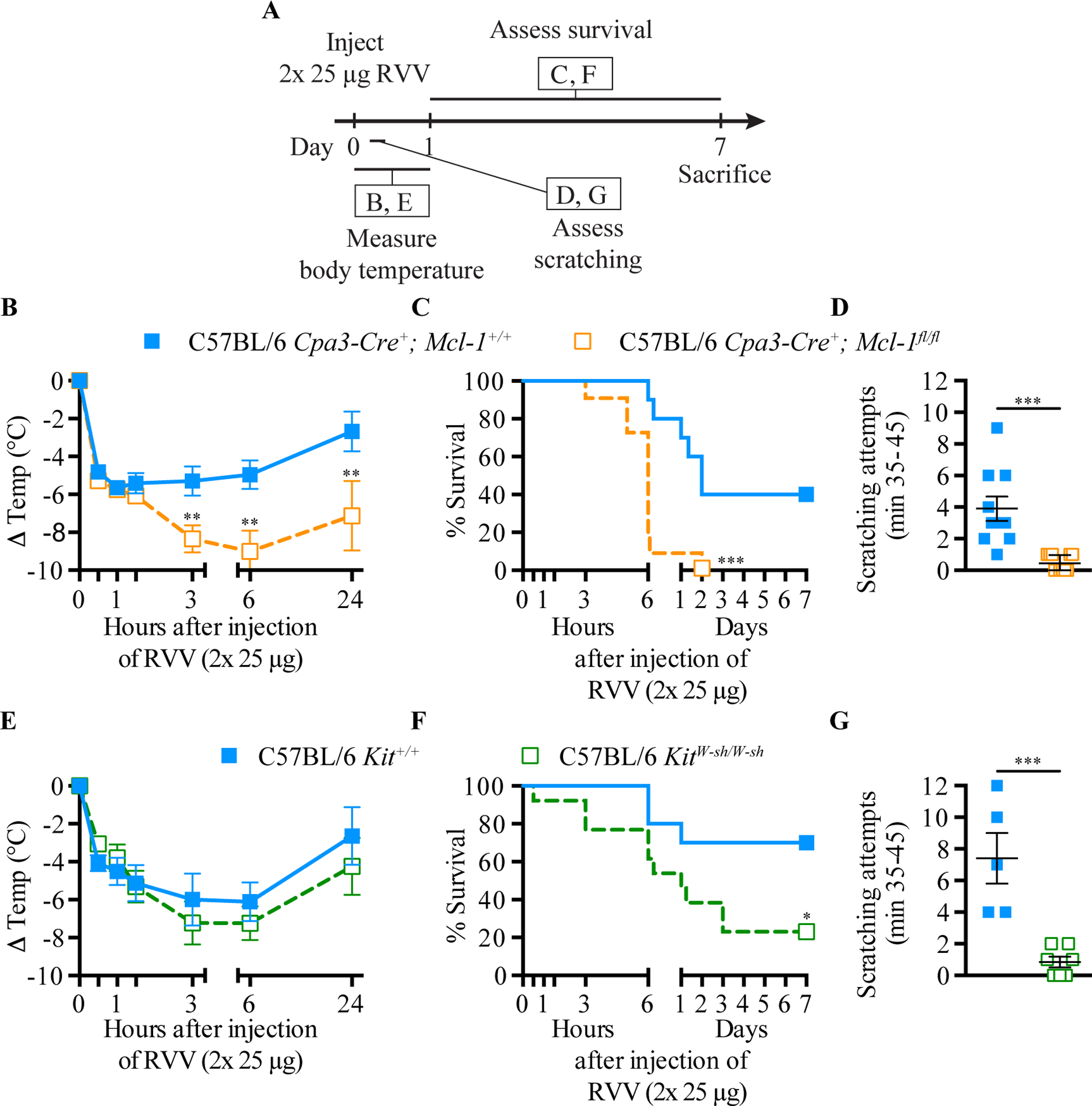
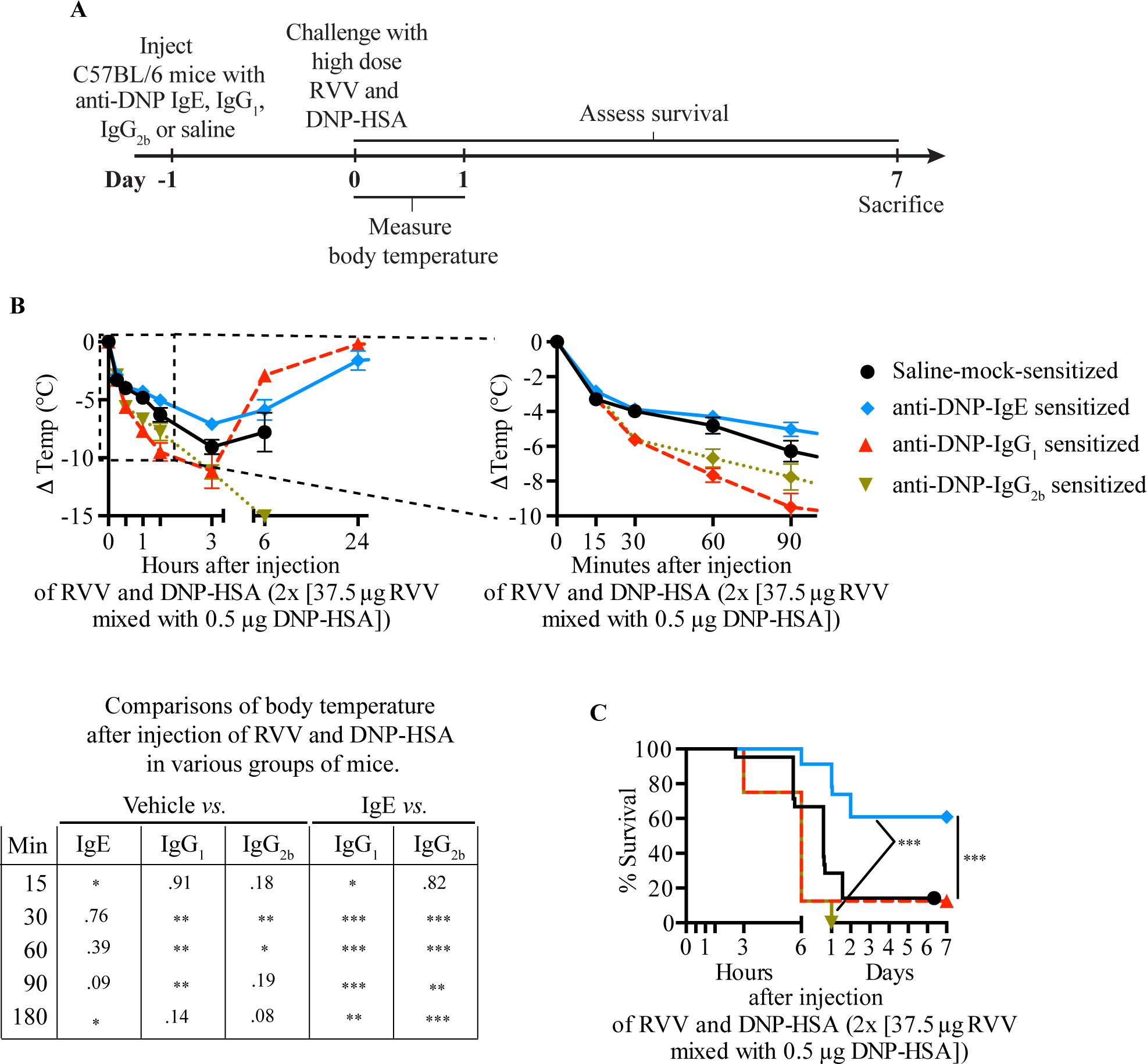
Physicians think of mast cells and IgE primarily in the context of allergic disorders, including fatal anaphylaxis. This 'bad side' of mast cells and IgE is so well accepted that it can be difficult to think of them in other contexts, particularly those in which they may have beneficial functions. However, there is evidence that mast cells and IgE, as well as basophils (circulating granulocytes whose functions partially overlap with those of mast cells), can contribute to host defense as components of adaptive type 2 immune responses to helminths, ticks and certain other parasites. Accordingly, allergies often are conceptualized as "misdirected" type 2 immune responses, in which IgE antibodies are produced against any of a diverse group of apparently harmless antigens, and against components of animal venoms. Indeed, certain unfortunate patients who have become sensitized to venoms develop severe IgE-associated allergic reactions, including fatal anaphylaxis, upon subsequent venom exposure. In this review, we will describe evidence that mast cells can enhance innate resistance, and survival, to challenge with reptile or arthropod venoms during a first exposure to such venoms. We also will discuss findings indicating that, in mice surviving an initial encounter with venom, acquired type 2 immune responses, IgE antibodies, the high affinity IgE receptor (FcεRI), and mast cells can contribute to acquired resistance to the lethal effects of both honeybee venom and Russell's viper venom. These findings support the hypothesis that mast cells and IgE can help protect the host against venoms and perhaps other noxious substances.
Keywords: Allergy; IgE; Th2 cell immunity; toxin hypothesis; venom.
Conflict of interest statement
All authors declare no conflict of interest.
Figures
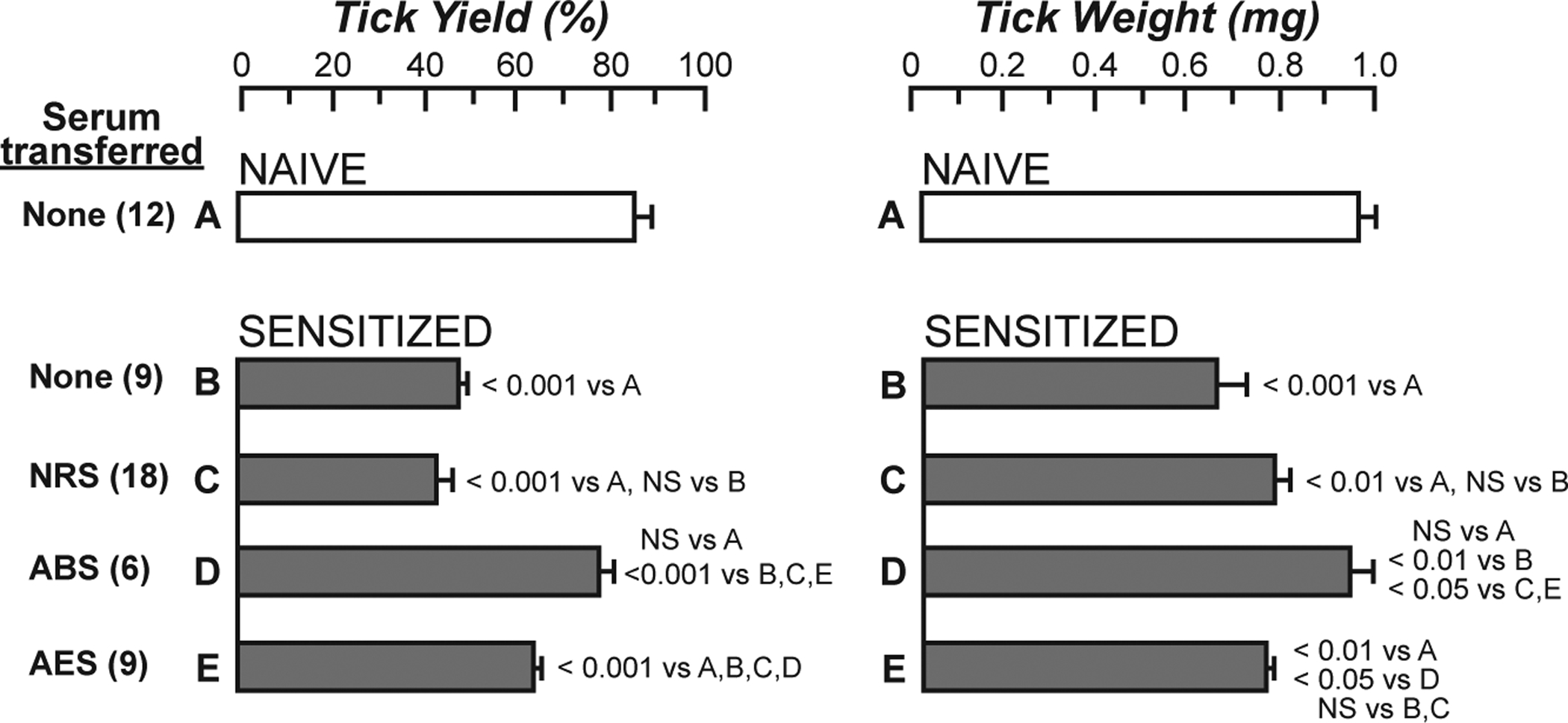
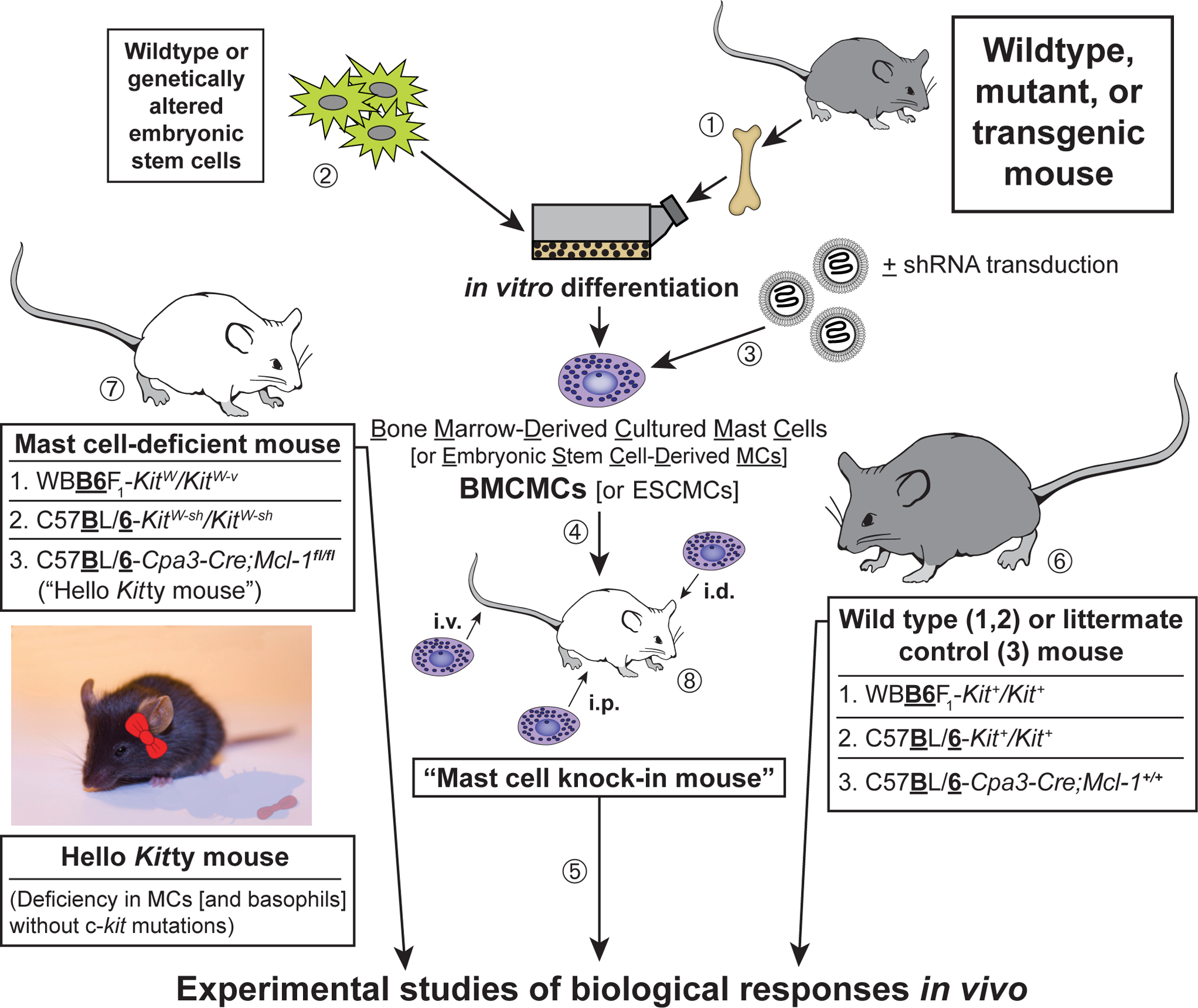
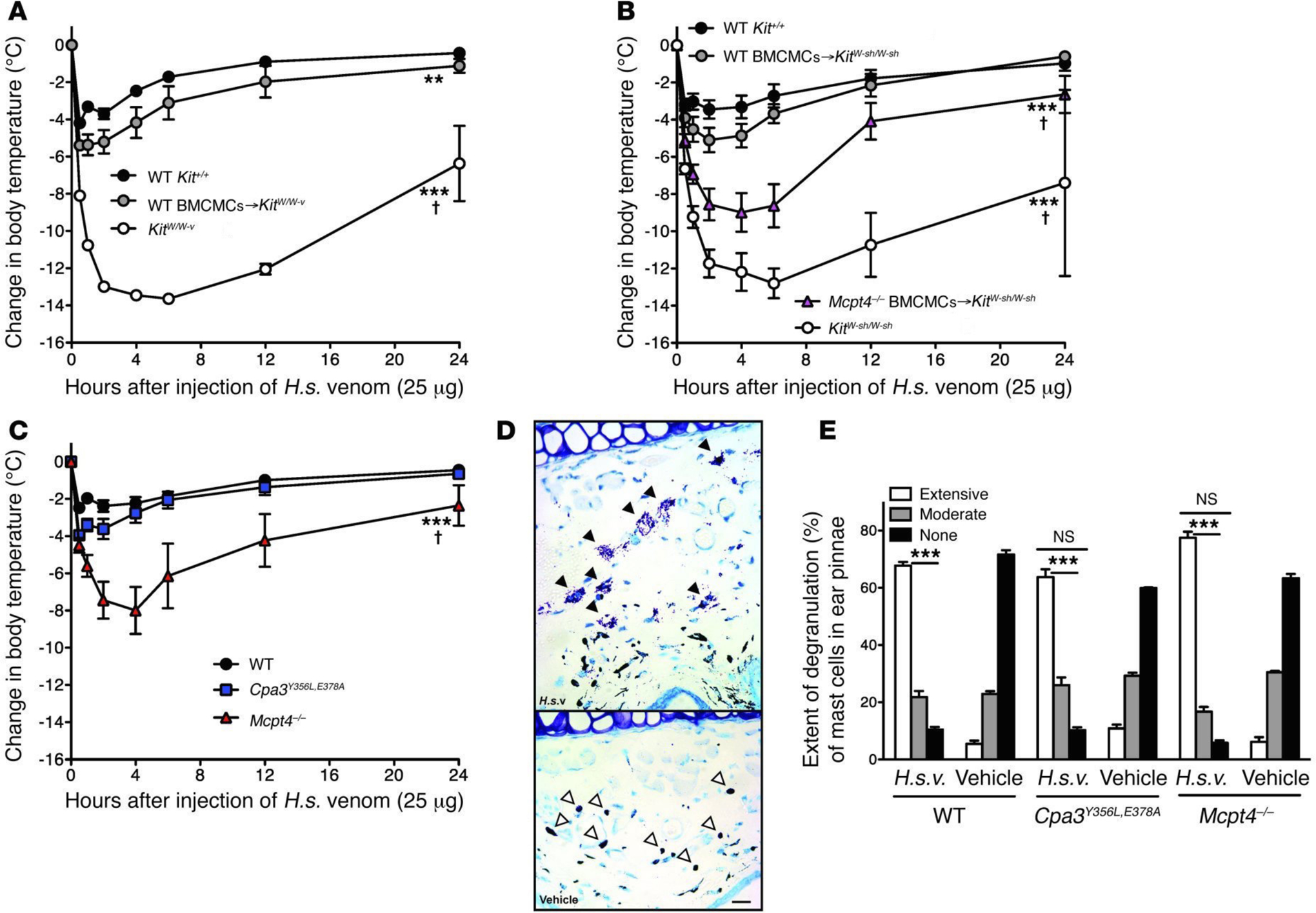
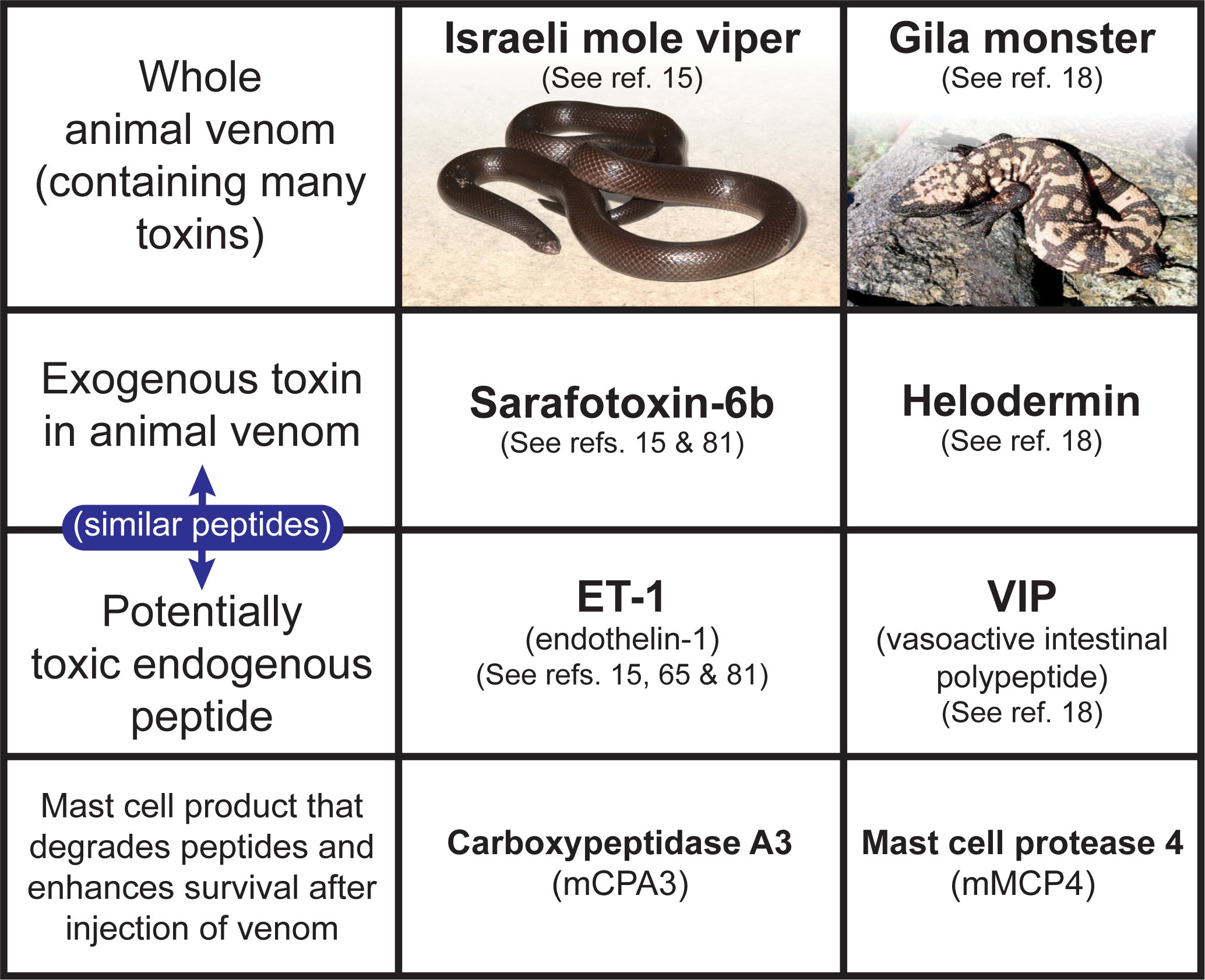
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
