

The power of randomization by sex in multilocus genetic evolution
- PMID: 33225949
- PMCID: PMC7682110
- DOI: 10.1186/s13062-020-00277-0
The power of randomization by sex in multilocus genetic evolution
Abstract
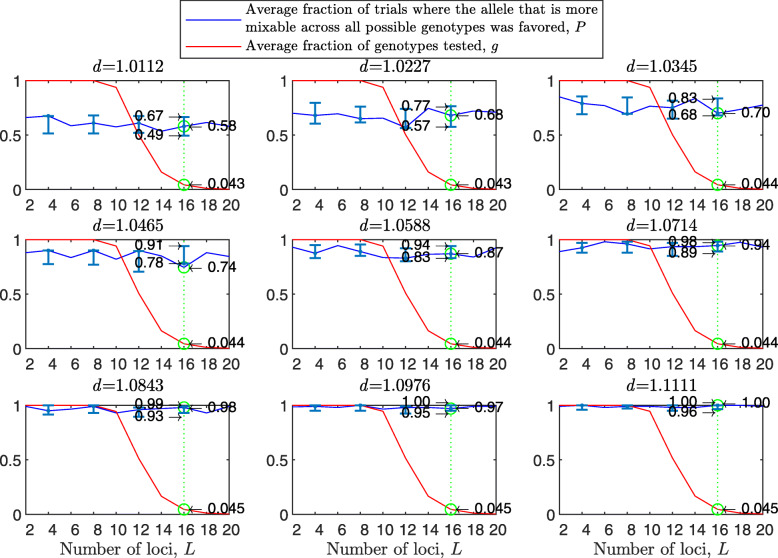
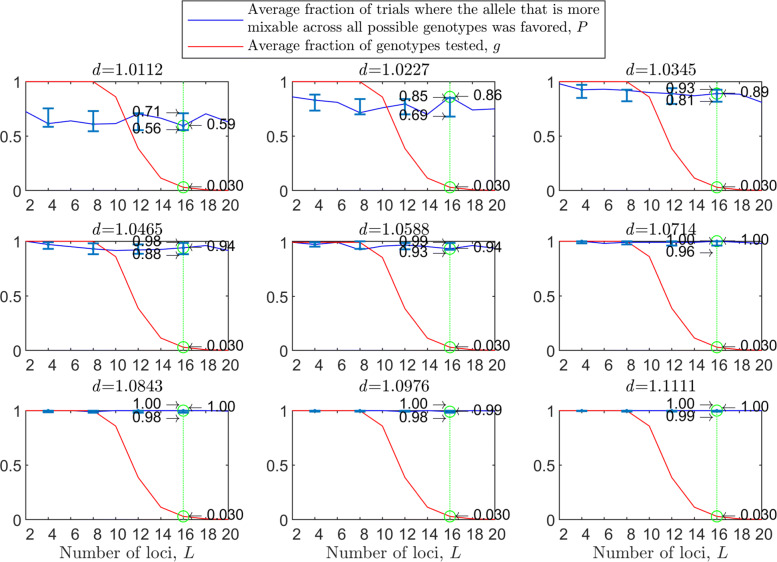
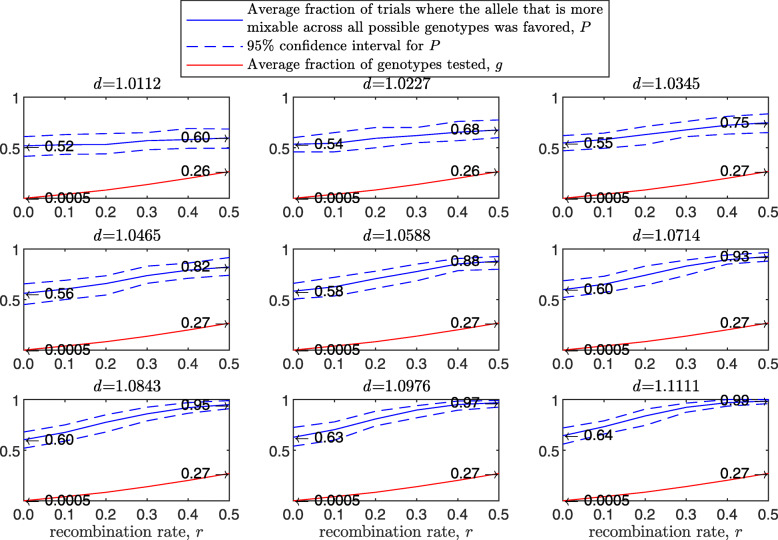
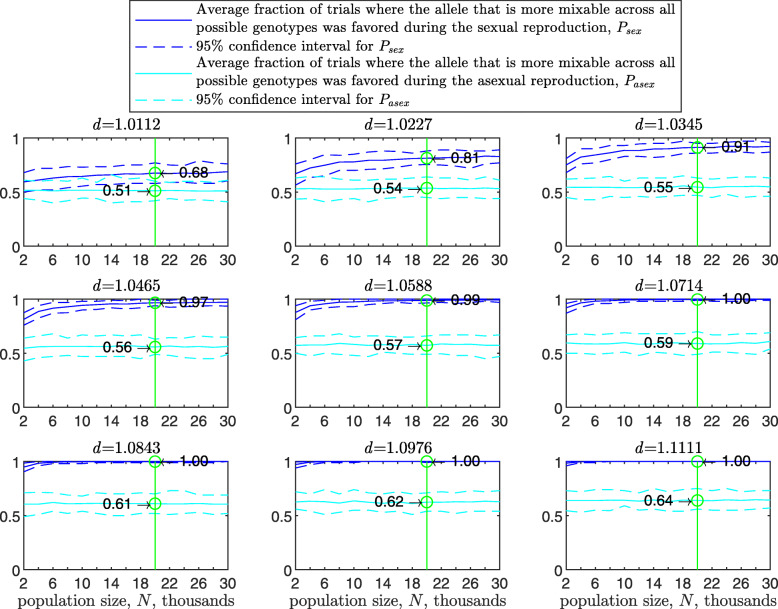
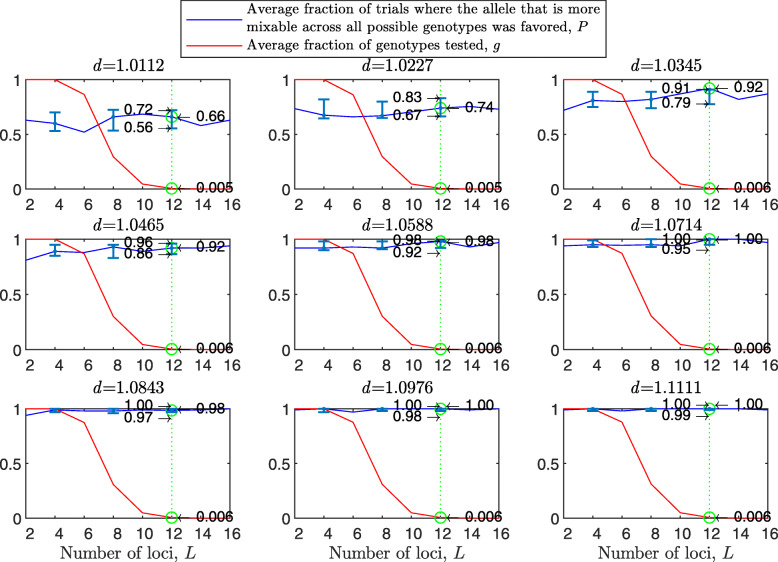
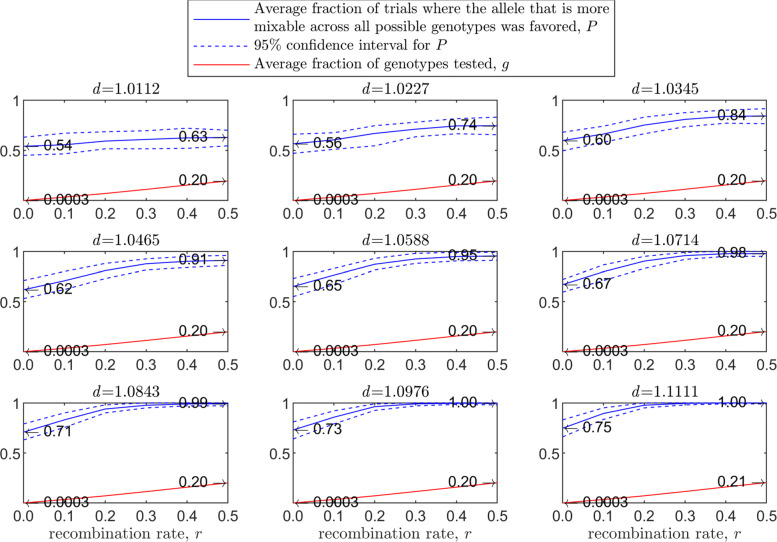
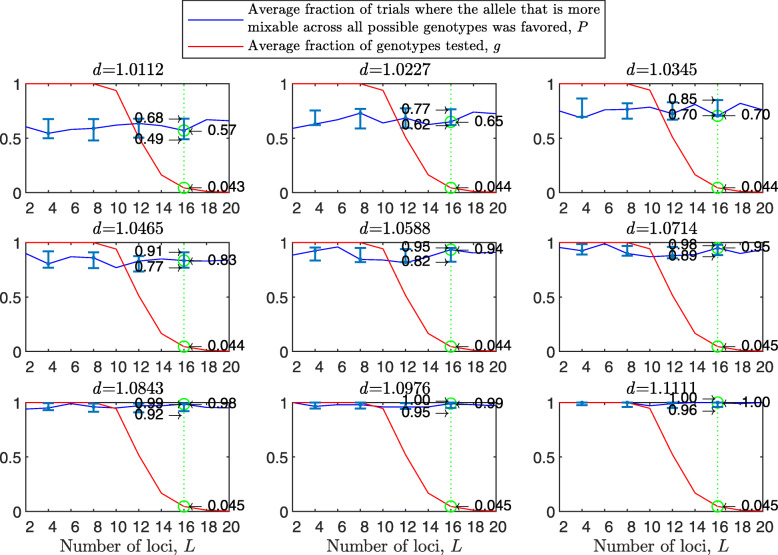
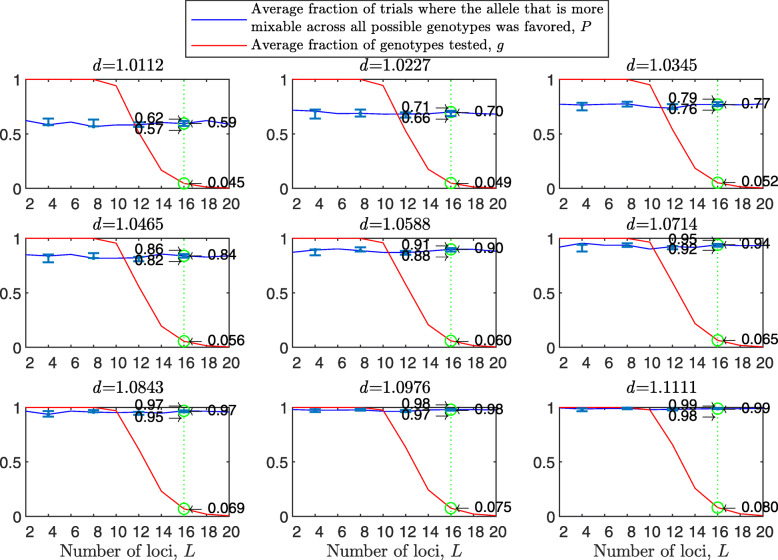
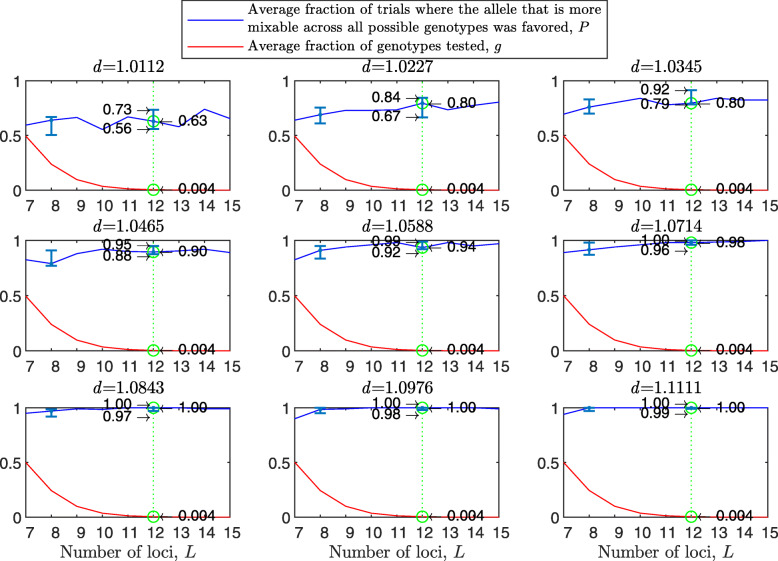
Background: Many hypotheses have been proposed for how sexual reproduction may facilitate an increase in the population mean fitness, such as the Fisher-Muller theory, Muller's ratchet and others. According to the recently proposed mixability theory, however, sexual recombination shifts the focus of natural selection away from favoring particular genetic combinations of high fitness towards favoring alleles that perform well across different genetic combinations. Mixability theory shows that, in finite populations, because sex essentially randomizes genetic combinations, if one allele performs better than another across the existing combinations of alleles, that allele will likely also perform better overall across a vast space of untested potential genotypes. However, this superiority has been established only for a single-locus diploid model.
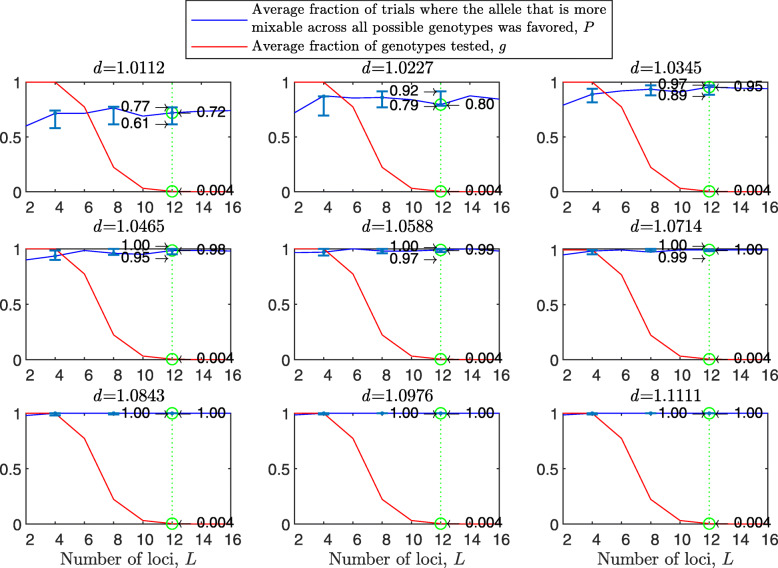
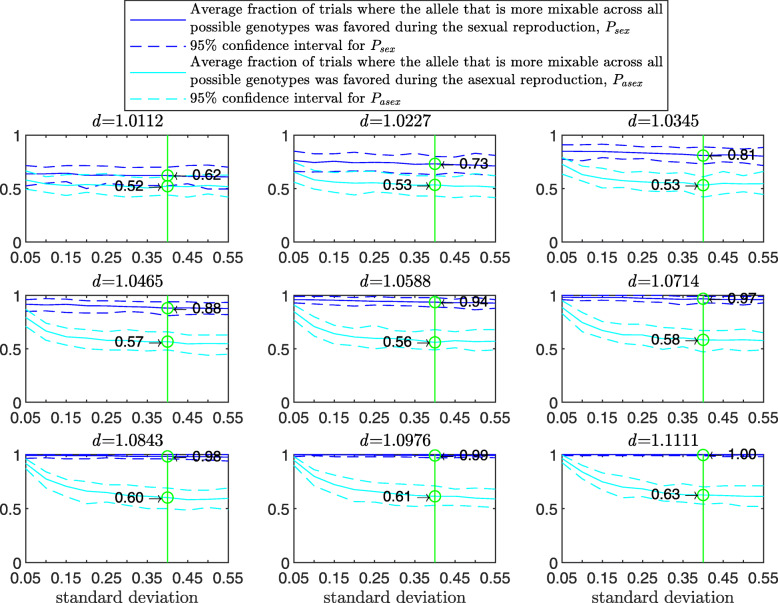
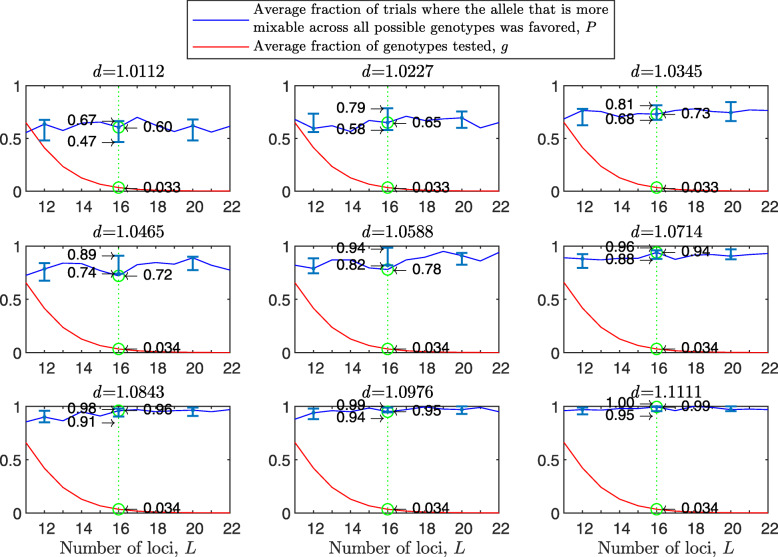
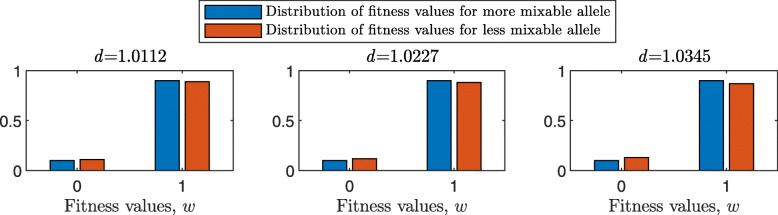
Results: We show that, in both haploids and diploids, the power of randomization by sex extends to the multilocus case, and becomes substantially stronger with increasing numbers of loci. In addition, we make an explicit comparison between the sexual and asexual cases, showing that sexual recombination is the cause of the randomization effect.
Conclusions: That the randomization effect applies to the multilocus case and becomes stronger with increasing numbers of loci suggests that it holds under realistic conditions. One may expect, therefore, that in nature the ability of an allele to perform well in interaction with existing genetic combinations is indicative of how well it will perform in a far larger space of potential combinations that have not yet materialized and been tested. Randomization plays a similar role in a statistical test, where it allows one to draw an inference from the outcome of the test in a small sample about its expected outcome in a larger space of possibilities-i.e., to generalize. Our results are relevant to recent theories examining evolution as a learning process.
Reviewers: This article was reviewed by David Ardell and Brian Golding.
Keywords: Epistasis; Interaction-based evolution; Multilocus models; Random sampling; Randomized algorithms; Sex and recombination.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures







Similar articles
-
The advantage of recombination when selection is acting at many genetic Loci.J Theor Biol. 2018 Apr 7;442:123-128. doi: 10.1016/j.jtbi.2018.01.018. J Theor Biol. 2018. PMID: 29355539
-
Sex: The power of randomization.Theor Popul Biol. 2019 Oct;129:41-53. doi: 10.1016/j.tpb.2018.11.005. Epub 2019 Jan 9. Theor Popul Biol. 2019. PMID: 30638926
-
Relative effects of segregation and recombination on the evolution of sex in finite diploid populations.Heredity (Edinb). 2013 Dec;111(6):505-12. doi: 10.1038/hdy.2013.72. Epub 2013 Jul 31. Heredity (Edinb). 2013. PMID: 23900397 Free PMC article.
-
[Evolution of sex: role of deleterious mutation and mobile elements].Zh Obshch Biol. 2003 Nov-Dec;64(6):463-78. Zh Obshch Biol. 2003. PMID: 14723170 Review. Russian.
-
Evolution of sex and the molecular clock in RNA viruses.Gene. 1997 Dec 31;205(1-2):301-8. doi: 10.1016/s0378-1119(97)00405-8. Gene. 1997. PMID: 9461404 Review.
Cited by
-
Resampling the pool of genotypic possibilities: an adaptive function of sexual reproduction.BMC Ecol Evol. 2021 Jun 12;21(1):119. doi: 10.1186/s12862-021-01850-5. BMC Ecol Evol. 2021. PMID: 34118864 Free PMC article.
-
Global mapping of cancers: The Cancer Genome Atlas and beyond.Mol Oncol. 2021 Nov;15(11):2823-2840. doi: 10.1002/1878-0261.13056. Epub 2021 Jul 20. Mol Oncol. 2021. PMID: 34245122 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
