

KIR3DL3 Is an Inhibitory Receptor for HHLA2 that Mediates an Alternative Immunoinhibitory Pathway to PD1
- PMID: 33229411
- PMCID: PMC8284010
- DOI: 10.1158/2326-6066.CIR-20-0315
KIR3DL3 Is an Inhibitory Receptor for HHLA2 that Mediates an Alternative Immunoinhibitory Pathway to PD1
Abstract
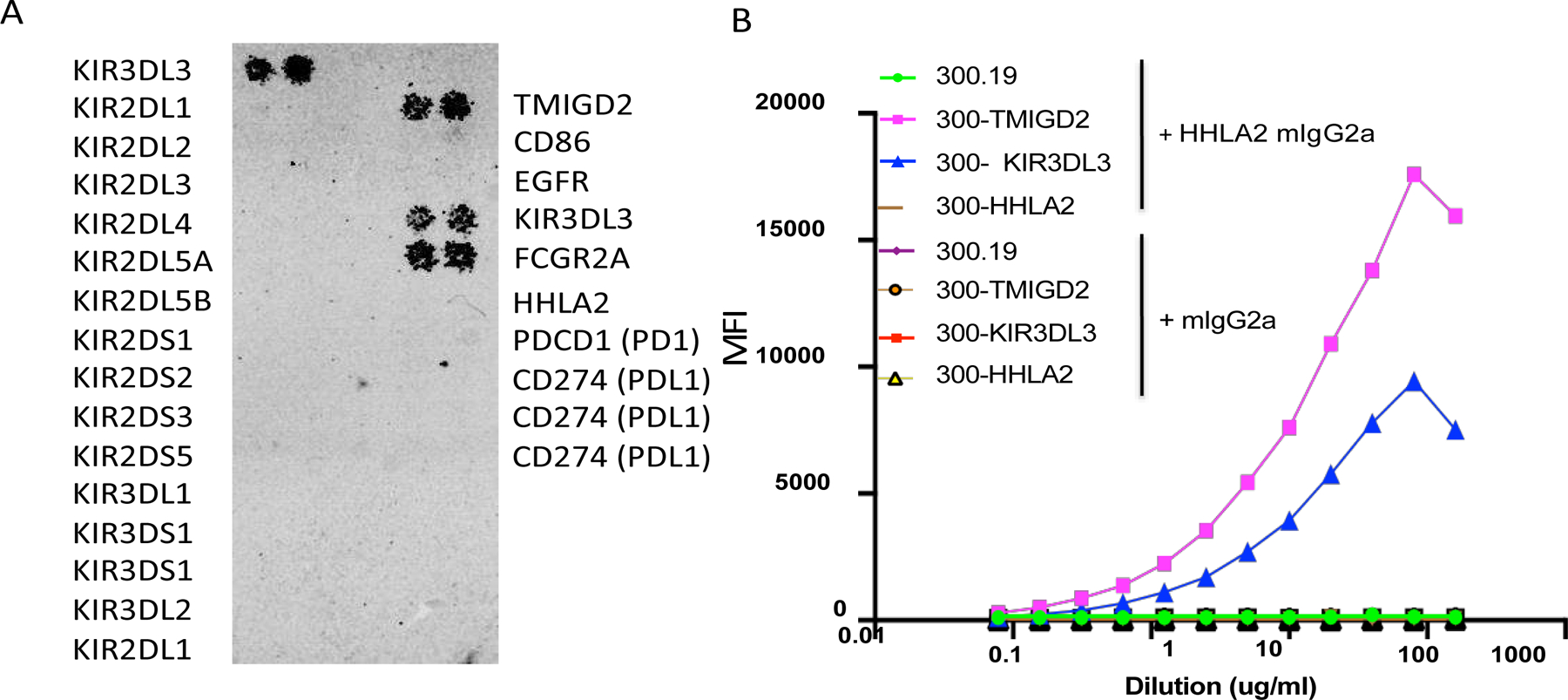
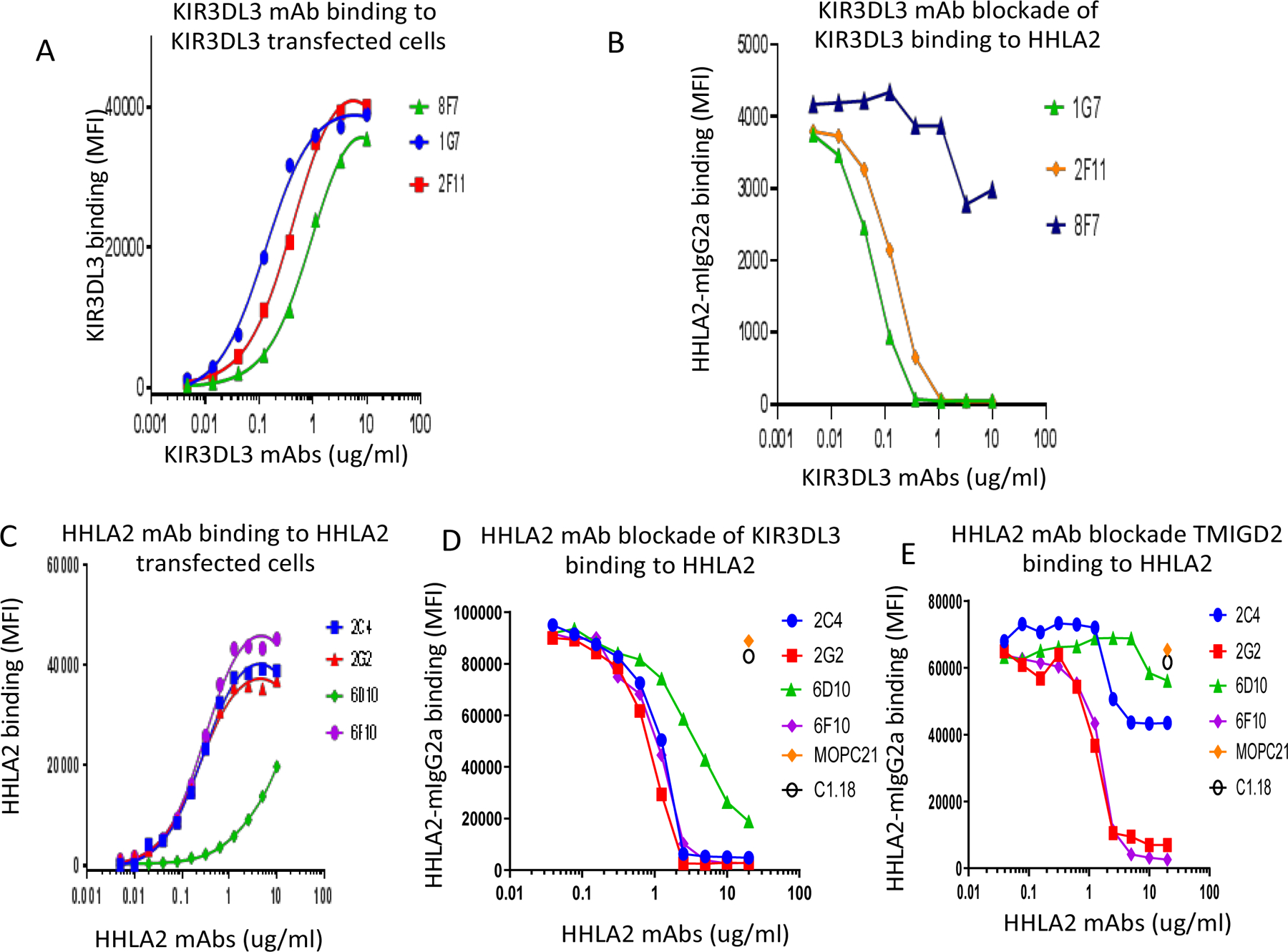
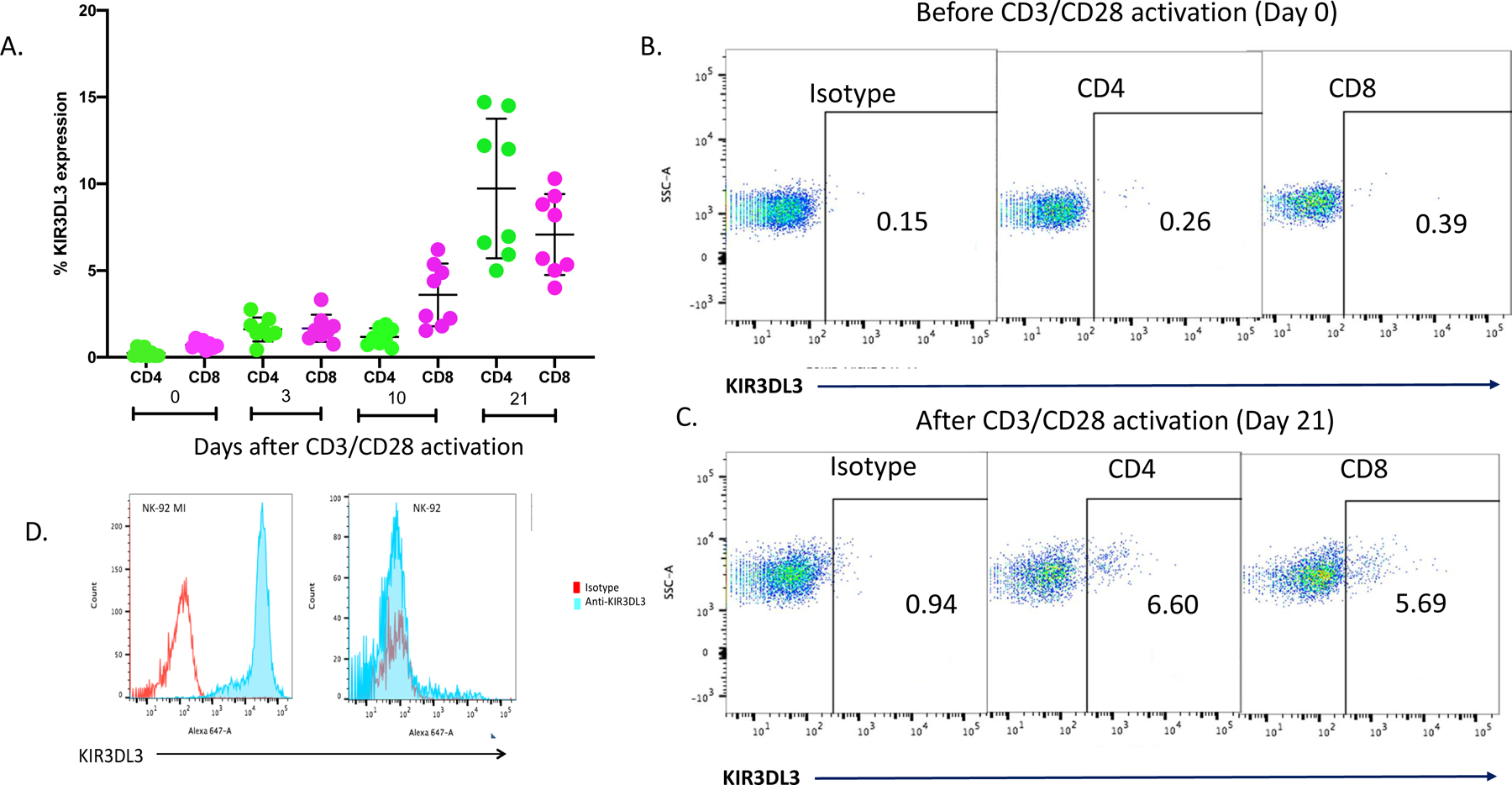
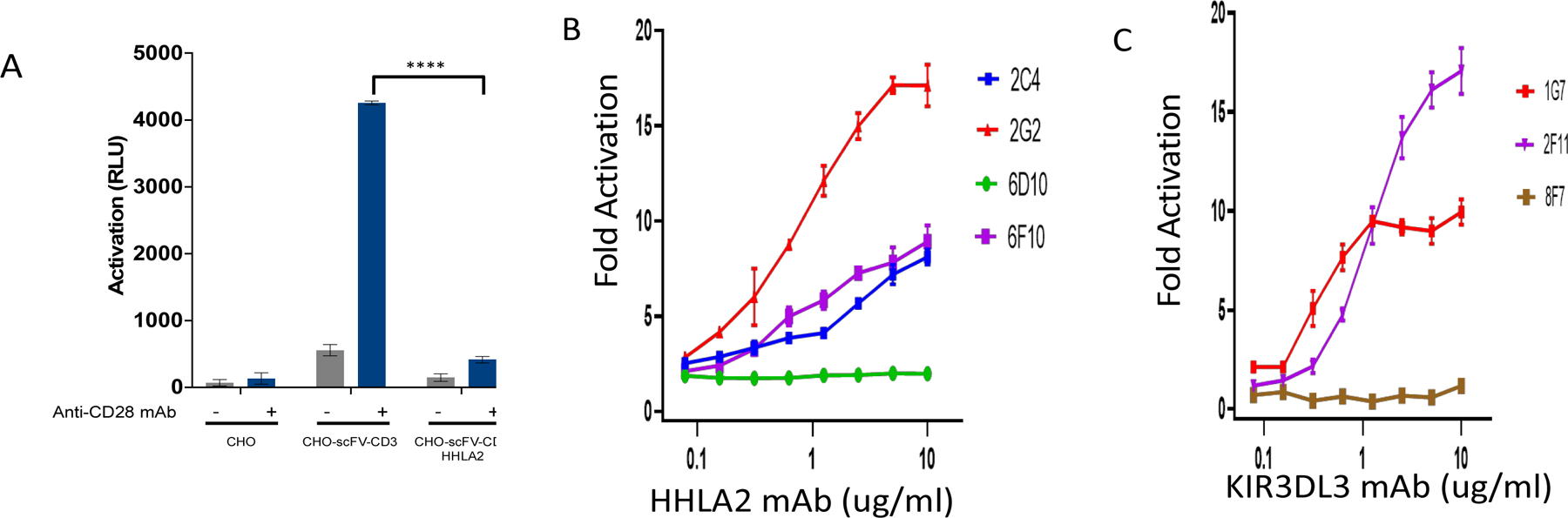
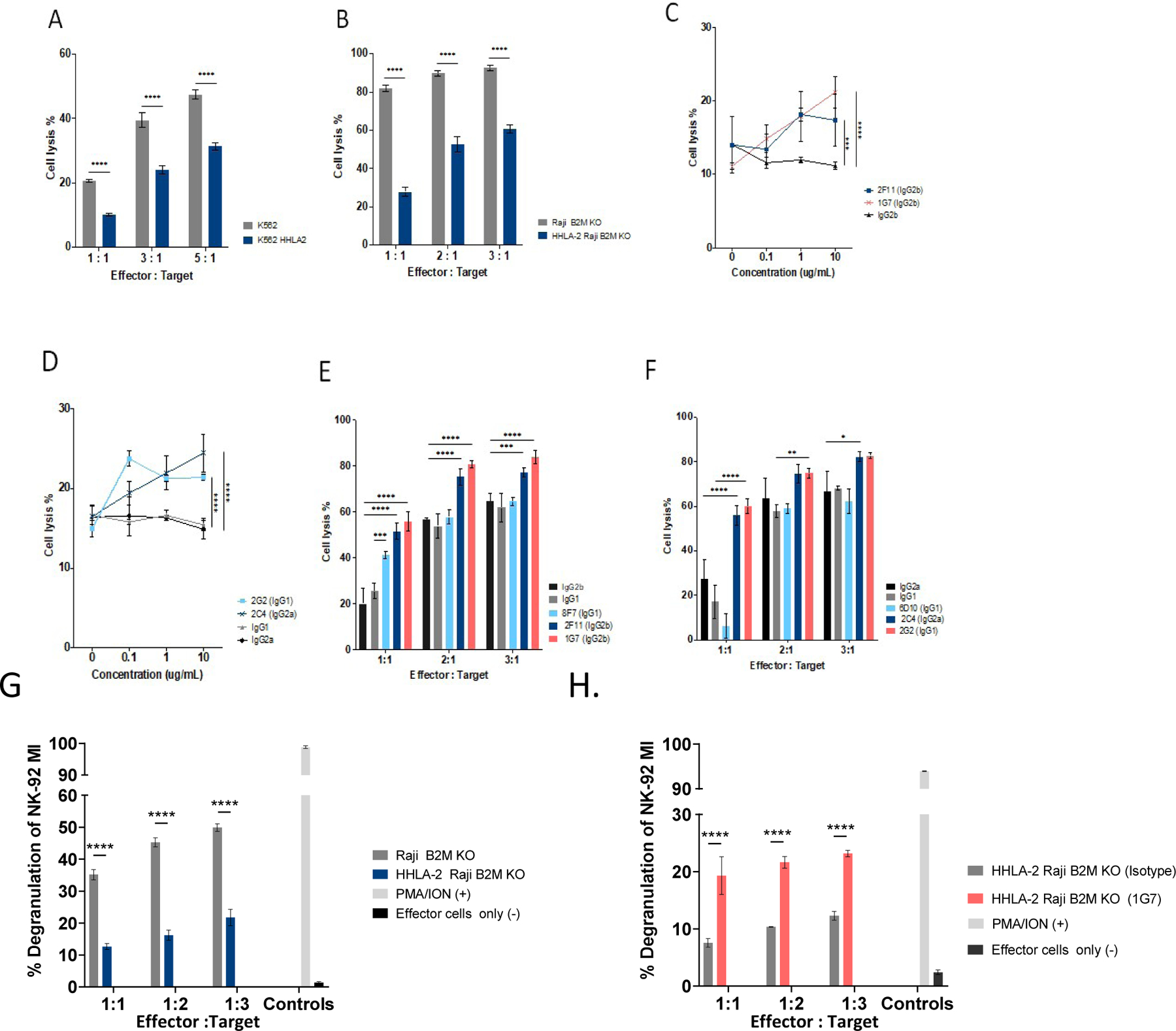
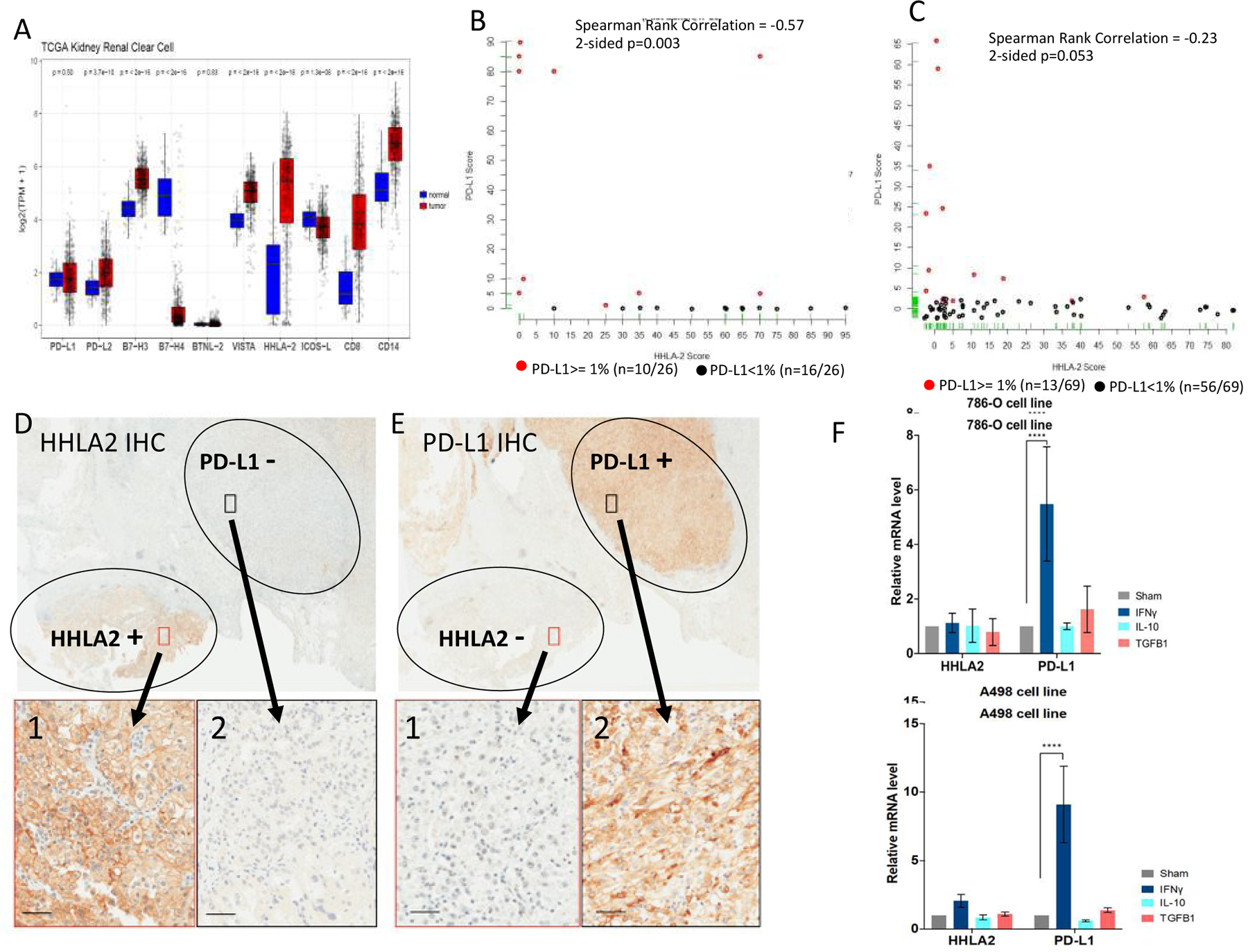
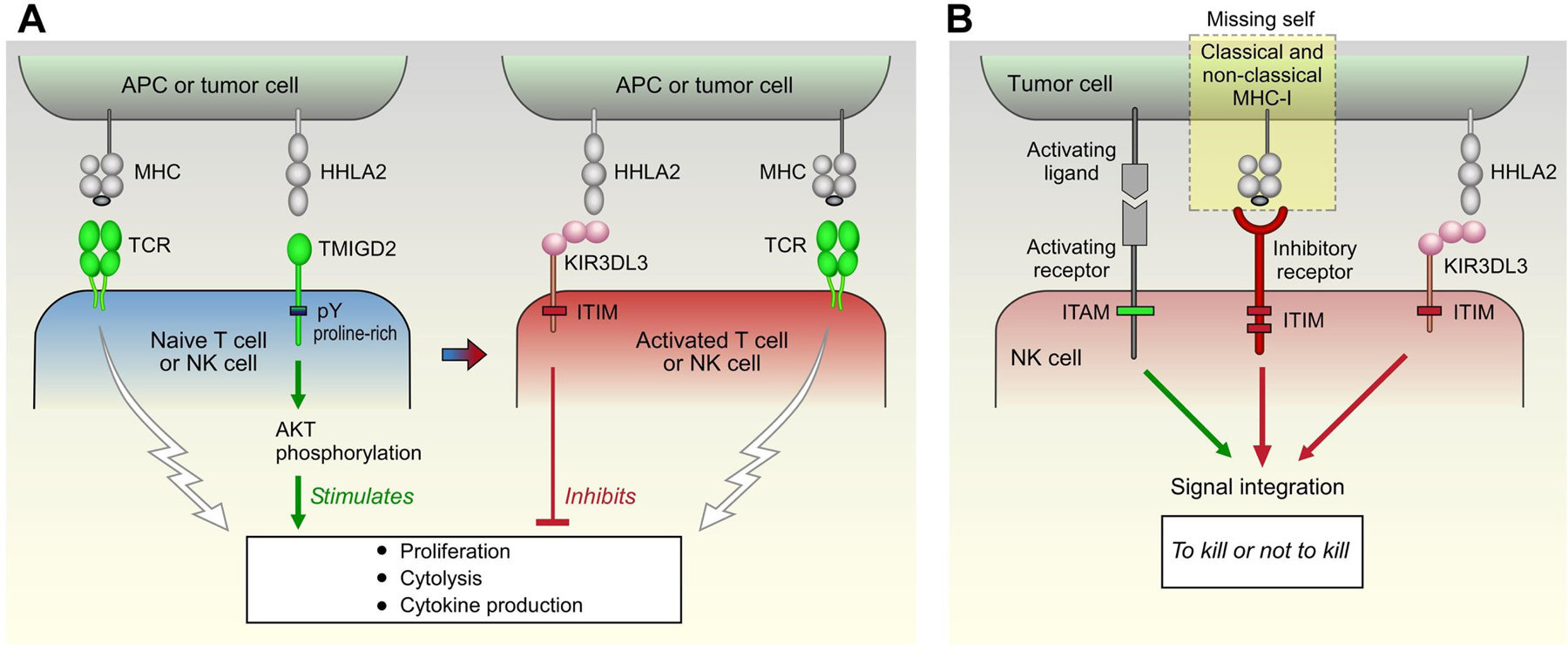
Blockade of the PD1 pathway is a broadly effective cancer therapy, but additional immune-inhibitory pathways contribute to tumor immune evasion. HERV-H LTR-associating 2 (HHLA2; also known as B7H5 and B7H7) is a member of the B7 family of immunoregulatory ligands that mediates costimulatory effects through its interaction with the CD28 family member transmembrane and immunoglobulin domain containing 2 (TMIGD2). However, HHLA2 has also been known to have inhibitory effects on T cells. Here, we report that we have identified killer cell immunoglobulin-like receptor, three immunoglobulin domains and long cytoplasmic tail 3 (KIR3DL3) as an inhibitory receptor for HHLA2 in T cells and natural killer (NK) cells and have generated HHLA2 and KIR3DL3 antibodies that block the immune-inhibitory activity of HHLA2, preserving the costimulatory signal. It is known that HHLA2 is frequently expressed in several tumor types, including clear cell renal cell carcinoma (ccRCC). We found that HHLA2 expression was nonoverlapping with PDL1 expression in ccRCC, suggesting that HHLA2 mediates a mechanism of tumor immune evasion that is independent from PDL1. Blockade of both the PD1 and KIR3DL3 pathways may be a more effective way to reverse tumor immune evasion.See related Spotlight on p. 128.
©2020 American Association for Cancer Research.
Conflict of interest statement
Figures
Comment in
-
Mystery Checkpoint Revealed: KIR3DL3 Finally Found a Ligand in HHLA2.Cancer Immunol Res. 2021 Feb;9(2):128. doi: 10.1158/2326-6066.CIR-20-0996. Cancer Immunol Res. 2021. PMID: 33536266
References
-
- Baumeister SH, Freeman GJ, Dranoff G, Sharpe AH. Coinhibitory Pathways in Immunotherapy for Cancer. Annu Rev Immunol. 2016;34:539–73. - PubMed
-
- Sharma P, Allison JP. The future of immune checkpoint therapy. Science. 2015;348(6230):56–61. - PubMed
-
- Pitt JM, Vetizou M, Daillere R, Roberti MP, Yamazaki T, Routy B, et al. Resistance Mechanisms to Immune-Checkpoint Blockade in Cancer: Tumor-Intrinsic and -Extrinsic Factors. Immunity. 2016;44(6):1255–69. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
