

The mutualism effector MiSSP7 of Laccaria bicolor alters the interactions between the poplar JAZ6 protein and its associated proteins
- PMID: 33230111
- PMCID: PMC7683724
- DOI: 10.1038/s41598-020-76832-6
The mutualism effector MiSSP7 of Laccaria bicolor alters the interactions between the poplar JAZ6 protein and its associated proteins
Abstract
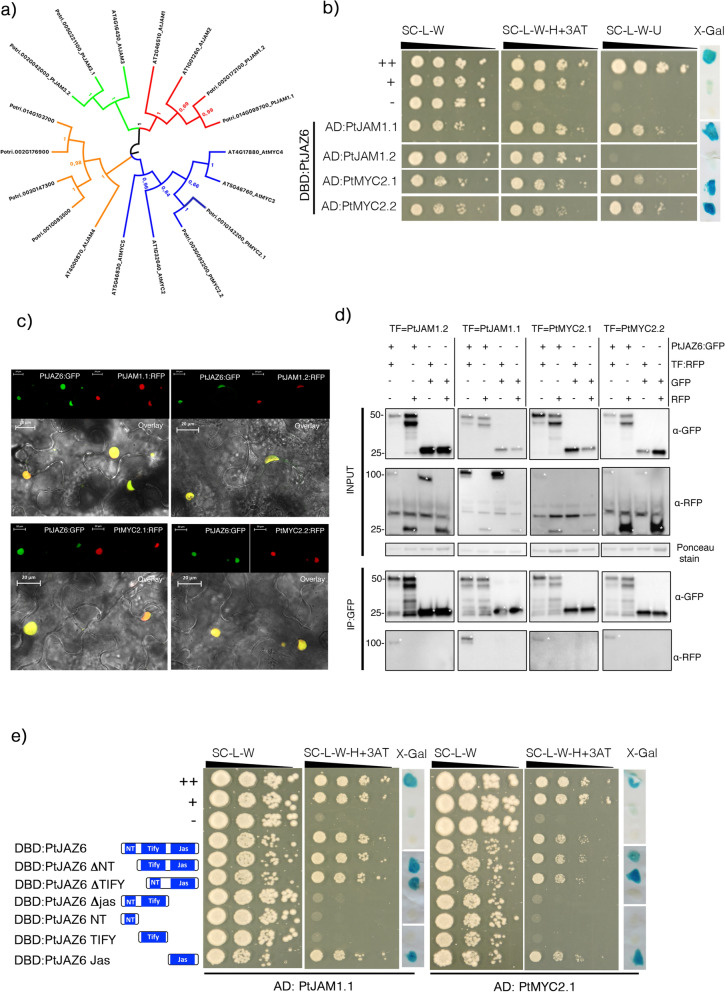
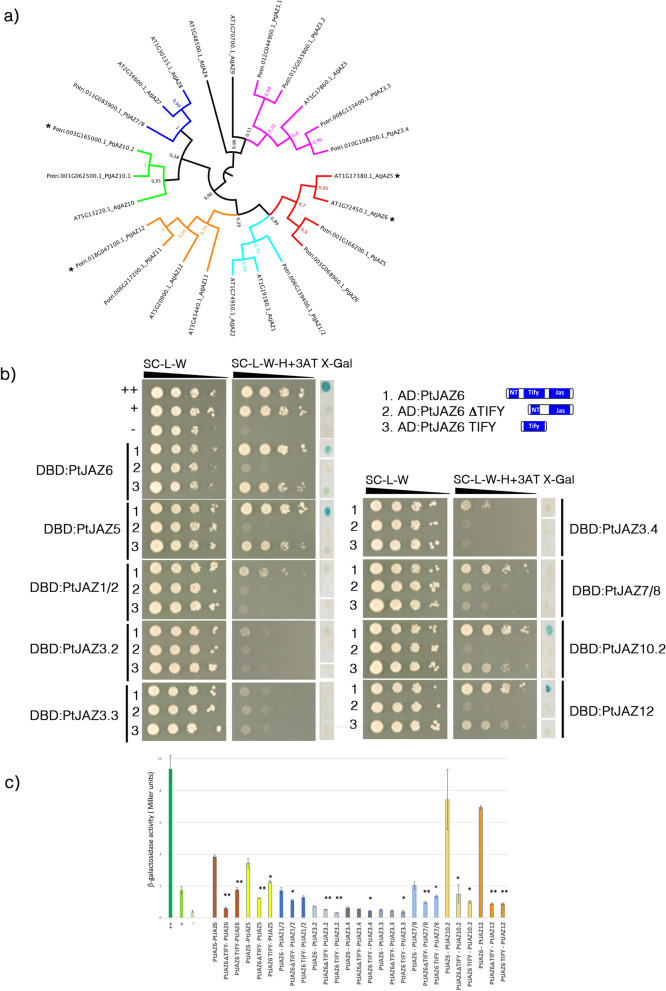
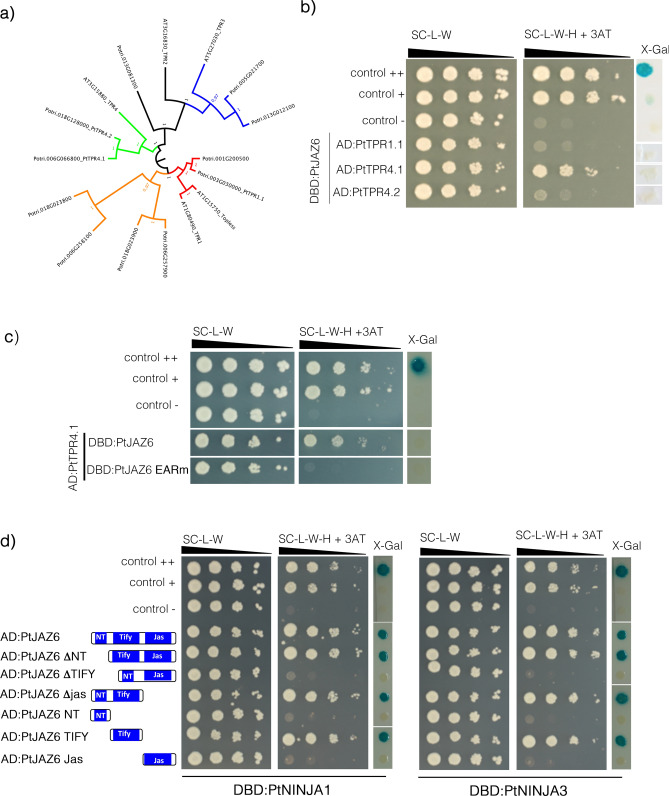
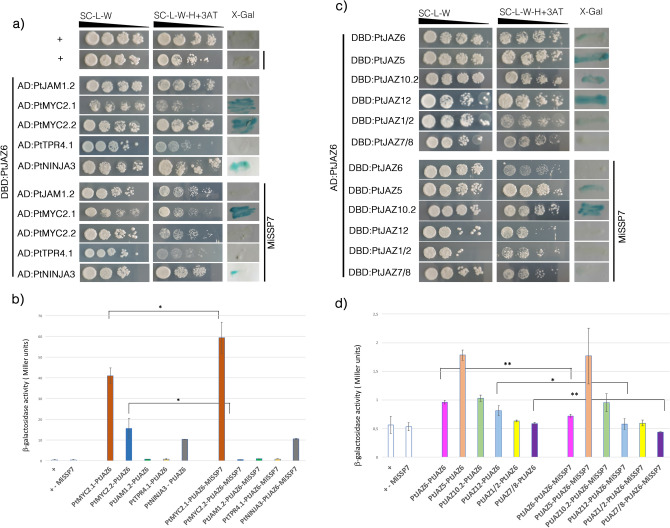
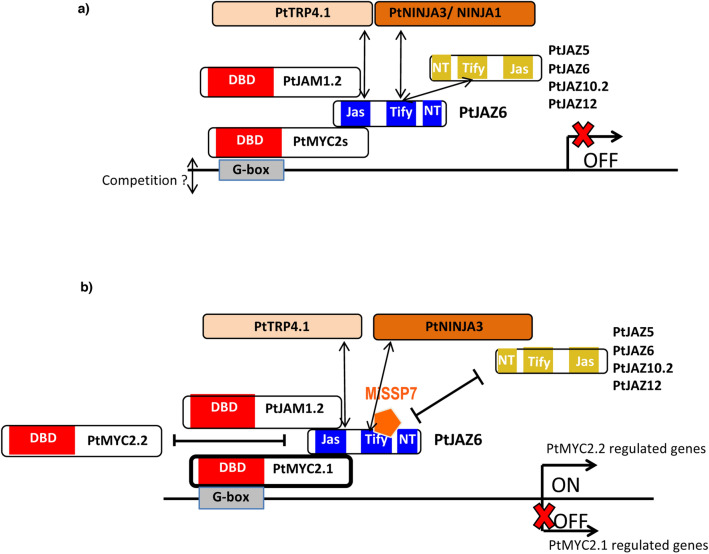
Despite the pivotal role of jasmonic acid in the outcome of plant-microorganism interactions, JA-signaling components in roots of perennial trees like western balsam poplar (Populus trichocarpa) are poorly characterized. Here we decipher the poplar-root JA-perception complex centered on PtJAZ6, a co-repressor of JA-signaling targeted by the effector protein MiSSP7 from the ectomycorrhizal basidiomycete Laccaria bicolor during symbiotic development. Through protein-protein interaction studies in yeast we determined the poplar root proteins interacting with PtJAZ6. Moreover, we assessed via yeast triple-hybrid how the mutualistic effector MiSSP7 reshapes the association between PtJAZ6 and its partner proteins. In the absence of the symbiotic effector, PtJAZ6 interacts with the transcription factors PtMYC2s and PtJAM1.1. In addition, PtJAZ6 interacts with it-self and with other Populus JAZ proteins. Finally, MiSSP7 strengthens the binding of PtJAZ6 to PtMYC2.1 and antagonizes PtJAZ6 homo-/heterodimerization. We conclude that a symbiotic effector secreted by a mutualistic fungus may promote the symbiotic interaction through altered dynamics of a JA-signaling-associated protein-protein interaction network, maintaining the repression of PtMYC2.1-regulated genes.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Bücking H, Hans R, Heyser W. The apoplast of ectomycorrhizal roots—site of nutrient uptake and nutrient exchange between the symbiotic partners. In: Sattelmacher B, Horst WJ, editors. The Apoplast of Higher Plants: Compartment of Storage, Transport and Reactions. Dordrecht: Springer; 2007. pp. 97–108.
-
- Gange AC, Gane DRJ, Chen YL, Gong MQ. Dual colonization of Eucalyptus urophylla ST Blake by arbuscular and ectomycorrhizal fungi affects levels of insect herbivore attack. Agric. For. Entomol. 2005;7:253–263. doi: 10.1111/j.1461-9555.2005.00268.x. - DOI
-
- Beniwal RS, Langenfeld-Heyser R, Polle A. Ectomycorrhiza and hydrogel protect hybrid poplar from water deficit and unravel plastic responses of xylem anatomy. Environ. Exp. Bot. 2010;69:189–197. doi: 10.1016/j.envexpbot.2010.02.005. - DOI
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
