

SUCLA2 mutations cause global protein succinylation contributing to the pathomechanism of a hereditary mitochondrial disease
- PMID: 33230181
- PMCID: PMC7684291
- DOI: 10.1038/s41467-020-19743-4
SUCLA2 mutations cause global protein succinylation contributing to the pathomechanism of a hereditary mitochondrial disease
Abstract
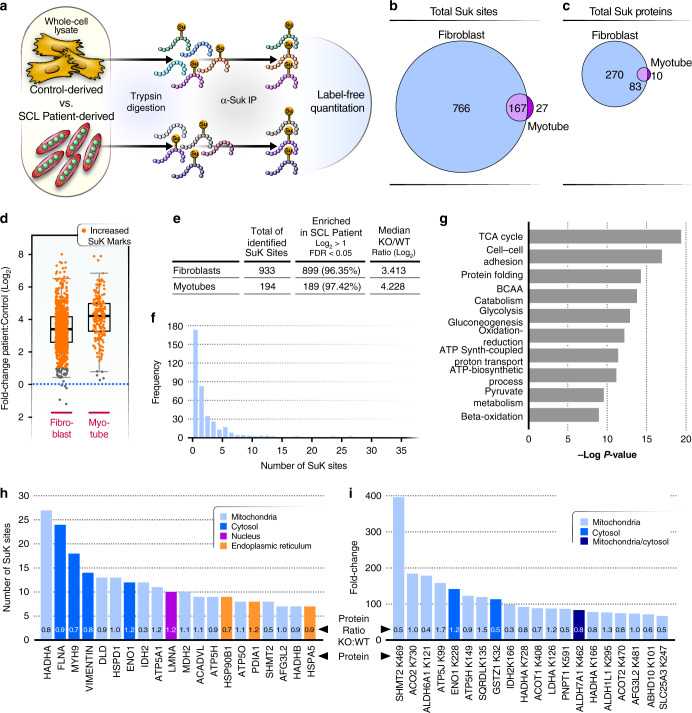
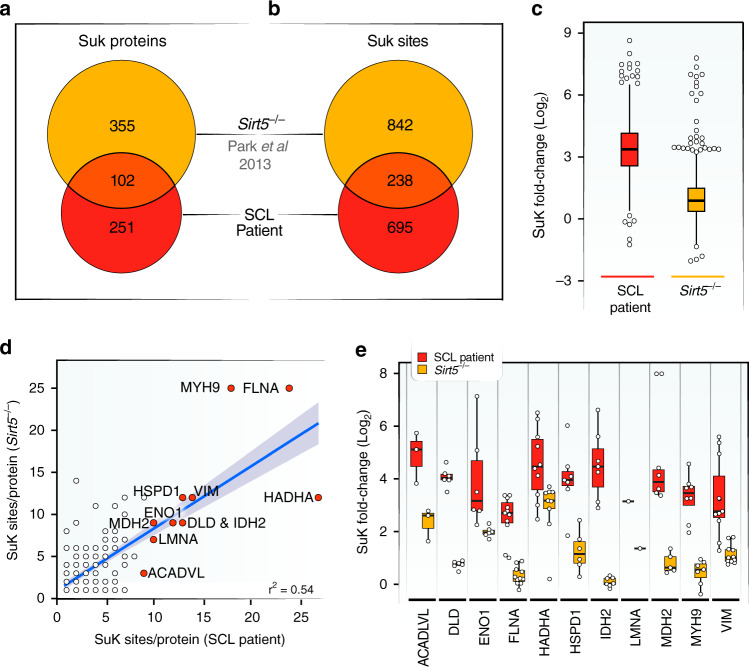
Mitochondrial acyl-coenzyme A species are emerging as important sources of protein modification and damage. Succinyl-CoA ligase (SCL) deficiency causes a mitochondrial encephalomyopathy of unknown pathomechanism. Here, we show that succinyl-CoA accumulates in cells derived from patients with recessive mutations in the tricarboxylic acid cycle (TCA) gene succinyl-CoA ligase subunit-β (SUCLA2), causing global protein hyper-succinylation. Using mass spectrometry, we quantify nearly 1,000 protein succinylation sites on 366 proteins from patient-derived fibroblasts and myotubes. Interestingly, hyper-succinylated proteins are distributed across cellular compartments, and many are known targets of the (NAD+)-dependent desuccinylase SIRT5. To test the contribution of hyper-succinylation to disease progression, we develop a zebrafish model of the SCL deficiency and find that SIRT5 gain-of-function reduces global protein succinylation and improves survival. Thus, increased succinyl-CoA levels contribute to the pathology of SCL deficiency through post-translational modifications.
Conflict of interest statement
J.R., G.C., J.T., S.C., S.M. and P.G. are employees of Nestlé Research. No patents have been filed related to work conducted in this study. The remaining authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
