

Limits to the cellular control of sequestered cryptophyte prey in the marine ciliate Mesodinium rubrum
- PMID: 33230263
- PMCID: PMC8115319
- DOI: 10.1038/s41396-020-00830-9
Limits to the cellular control of sequestered cryptophyte prey in the marine ciliate Mesodinium rubrum
Erratum in
-
Correction: Limits to the cellular control of sequestered cryptophyte prey in the marine ciliate Mesodinium rubrum.ISME J. 2022 Mar;16(3):898. doi: 10.1038/s41396-021-01115-5. ISME J. 2022. PMID: 34561549 Free PMC article. No abstract available.
Abstract
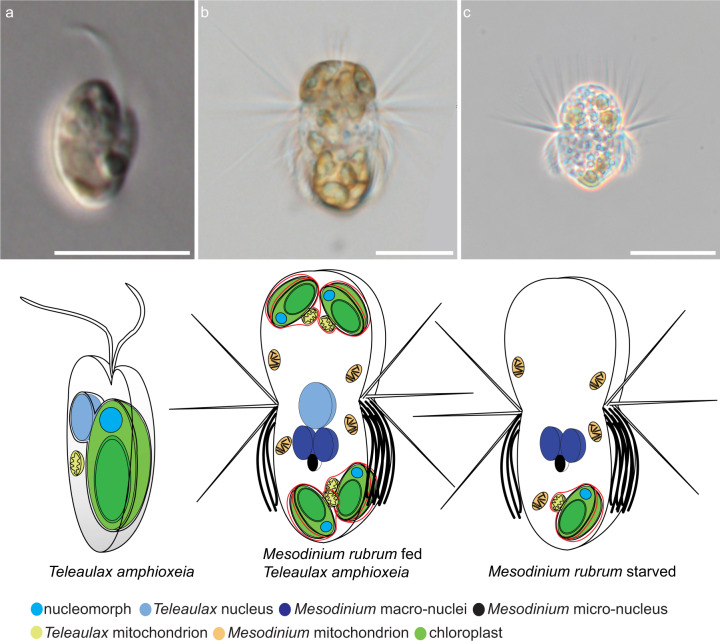
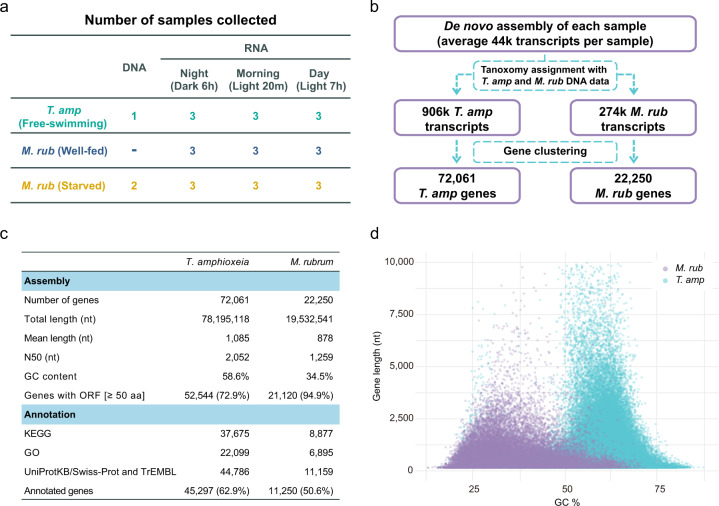
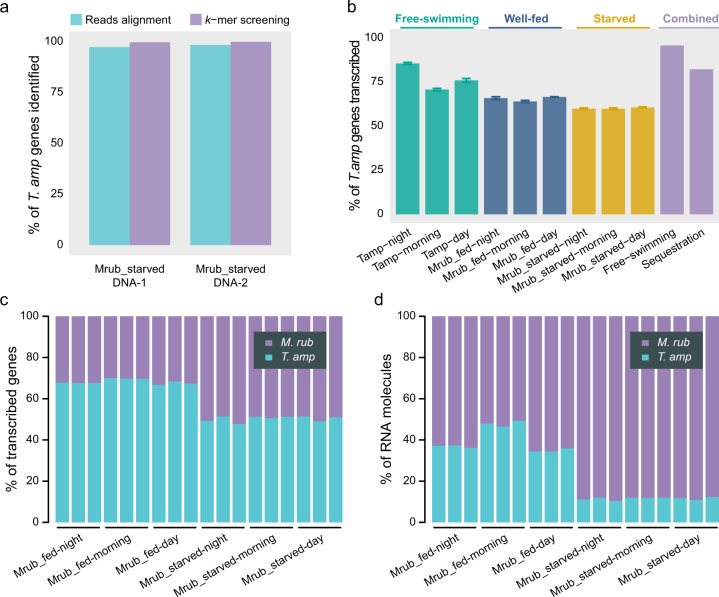
The marine ciliate Mesodinium rubrum is famous for its ability to acquire and exploit chloroplasts and other cell organelles from some cryptophyte algal species. We sequenced genomes and transcriptomes of free-swimming Teleaulax amphioxeia, as well as well-fed and starved M. rubrum in order to understand cellular processes upon sequestration under different prey and light conditions. From its prey, the ciliate acquires the ability to photosynthesize as well as the potential to metabolize several essential compounds including lysine, glycan, and vitamins that elucidate its specific prey dependency. M. rubrum does not express photosynthesis-related genes itself, but elicits considerable transcriptional control of the acquired cryptophyte organelles. This control is limited as light-dependent transcriptional changes found in free-swimming T. amphioxeia got lost after sequestration. We found strong transcriptional rewiring of the cryptophyte nucleus upon sequestration, where 35% of the T. amphioxeia genes were significantly differentially expressed within well-fed M. rubrum. Qualitatively, 68% of all genes expressed within well-fed M. rubrum originated from T. amphioxeia. Quantitatively, these genes contributed up to 48% to the global transcriptome in well-fed M. rubrum and down to 11% in starved M. rubrum. This tertiary endosymbiosis system functions for several weeks, when deprived of prey. After this point in time, the ciliate dies if not supplied with fresh prey cells. M. rubrum represents one evolutionary way of acquiring photosystems from its algal prey, and might represent a step on the evolutionary way towards a permanent tertiary endosymbiosis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
-
- Cavalier-Smith T. Membrane heredity and early chloroplast evolution. Trends Plant Sci. 2000;5:174–82. - PubMed
-
- Douglas SE, Murphy CA, Spencer DF, Gray MW. Cryptomonad algae are evolutionary chimaeras of two phylogenetically distinct unicellular eukaryotes. Nature. 1991;350:148–51. - PubMed
-
- Cavalier-Smith T. Principles of protein and lipid targeting in secondary symbiogenesis: euglenoid, dinoflagellate, and sporozoan plastid origins and the eukaryote family tree. J Eukaryot Microbiol. 1999;46:347–66. - PubMed
