

Auditory activity is diverse and widespread throughout the central brain of Drosophila
- PMID: 33230320
- PMCID: PMC7783861
- DOI: 10.1038/s41593-020-00743-y
Auditory activity is diverse and widespread throughout the central brain of Drosophila
Abstract
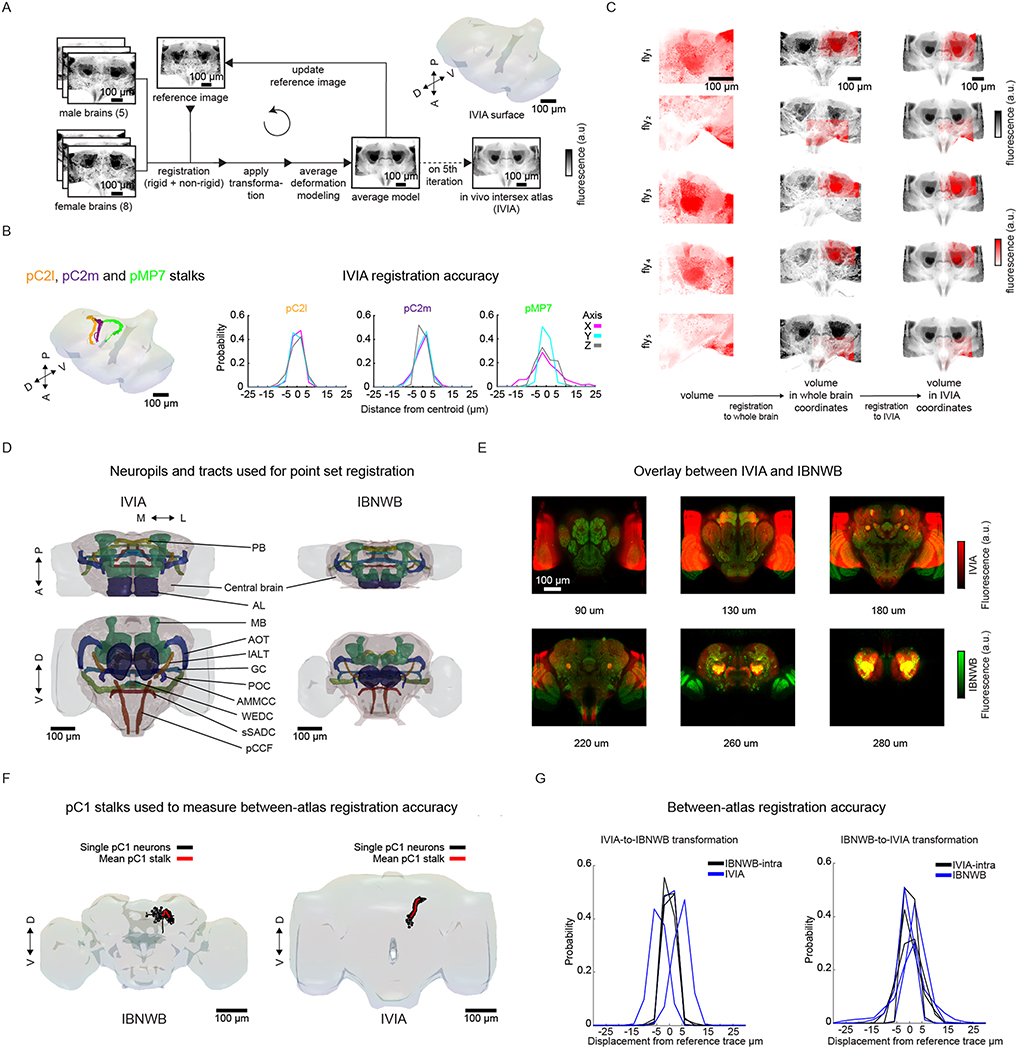
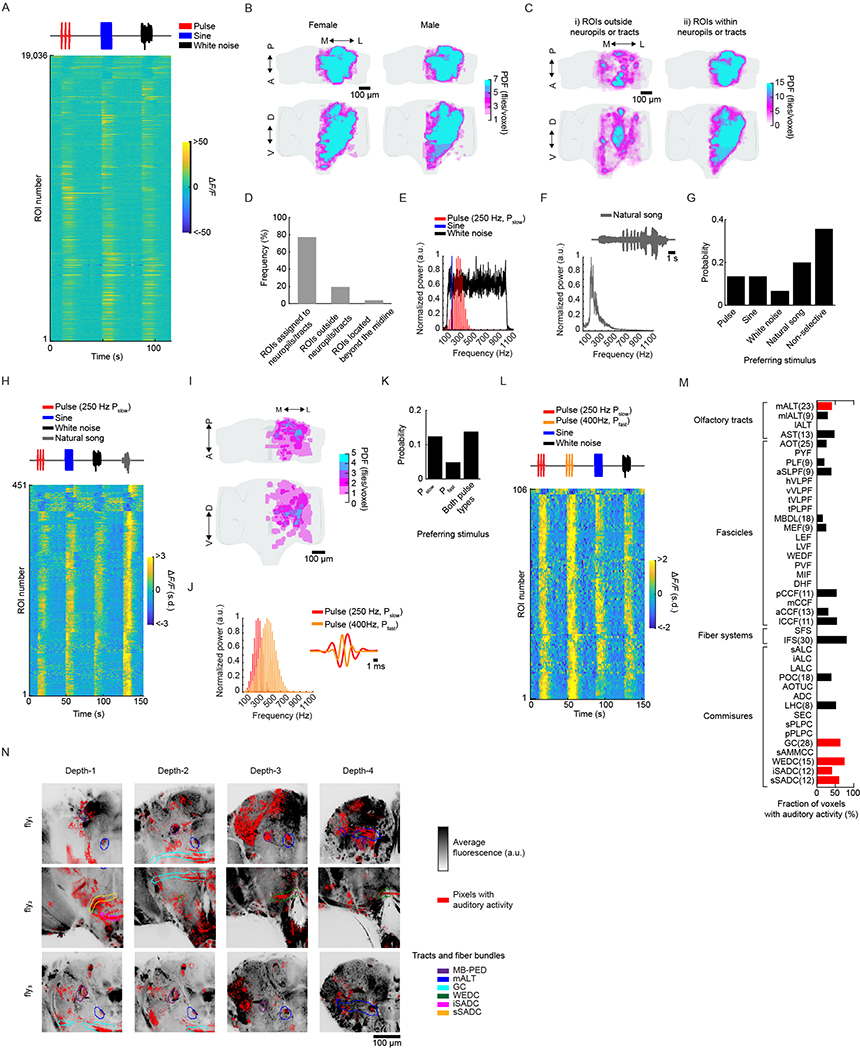
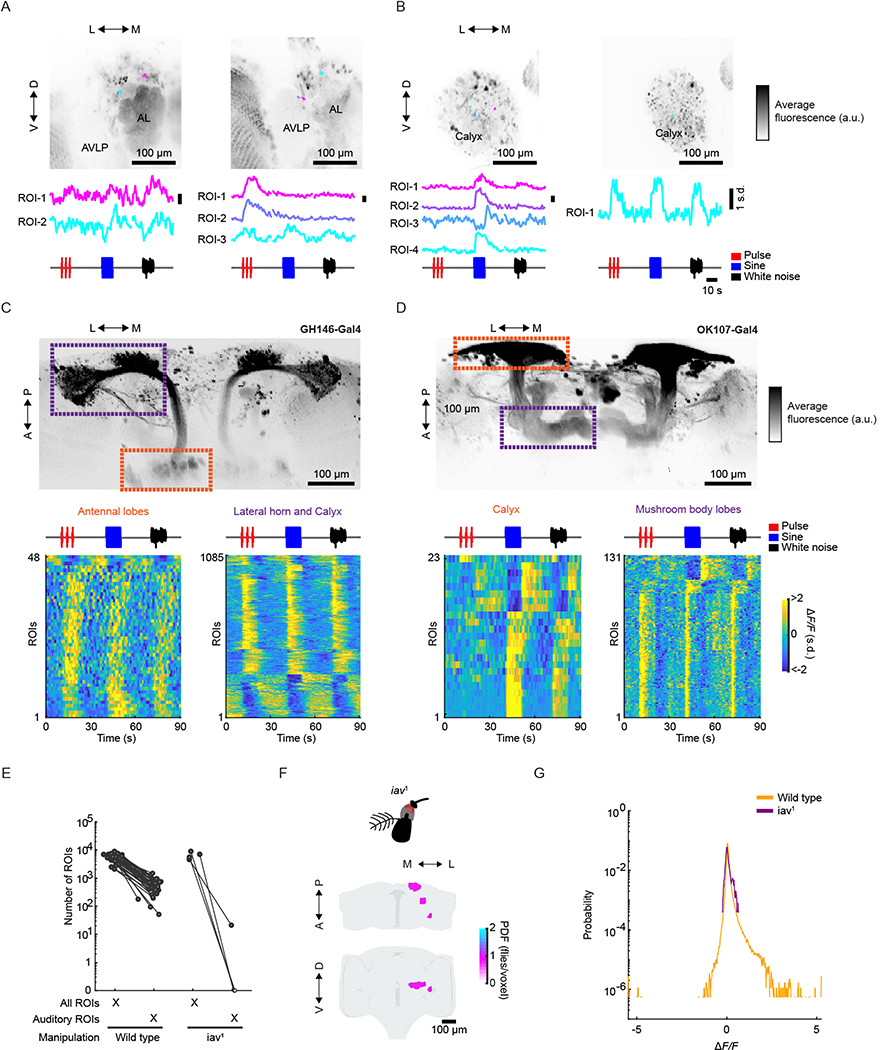
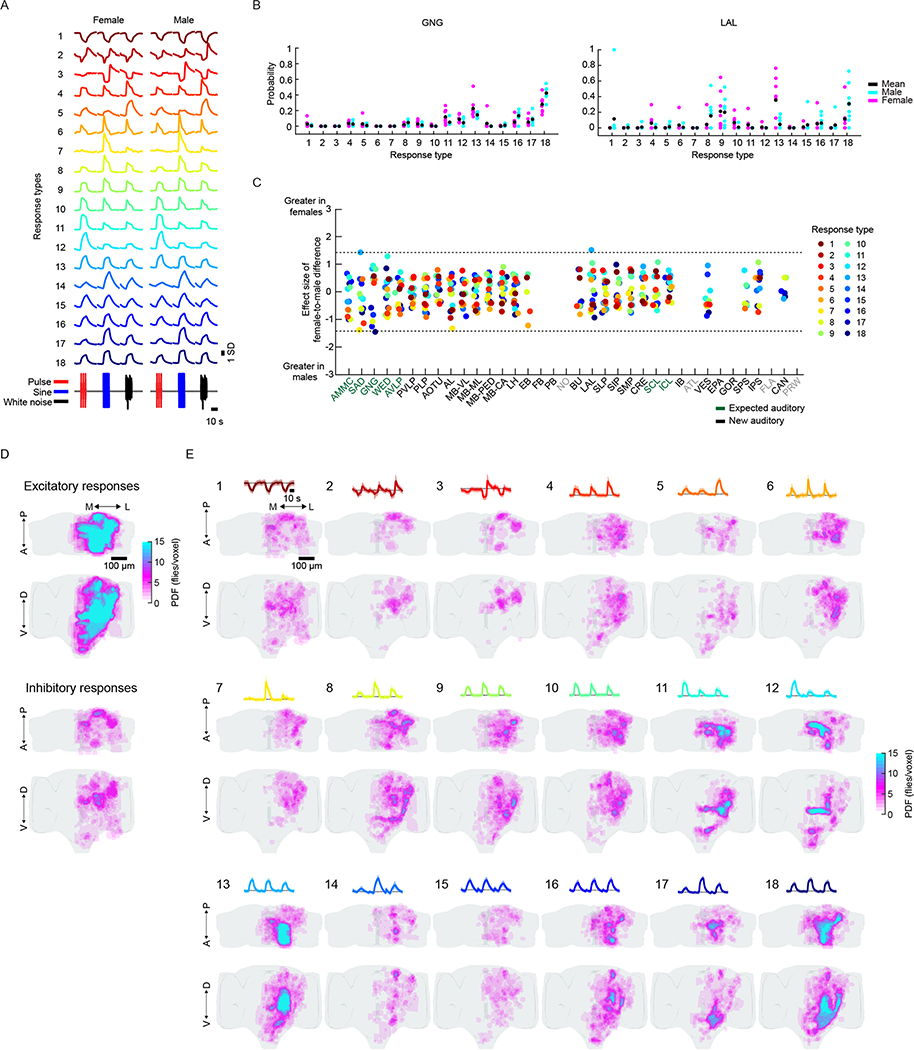
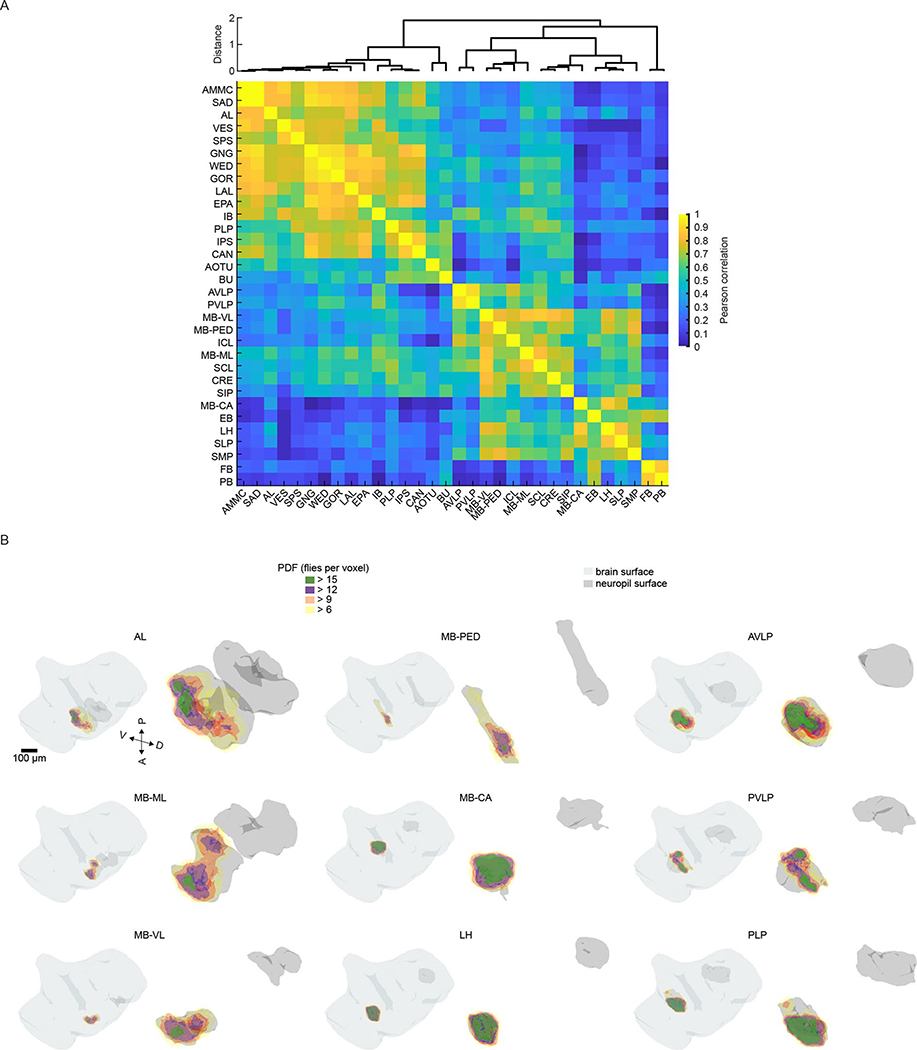
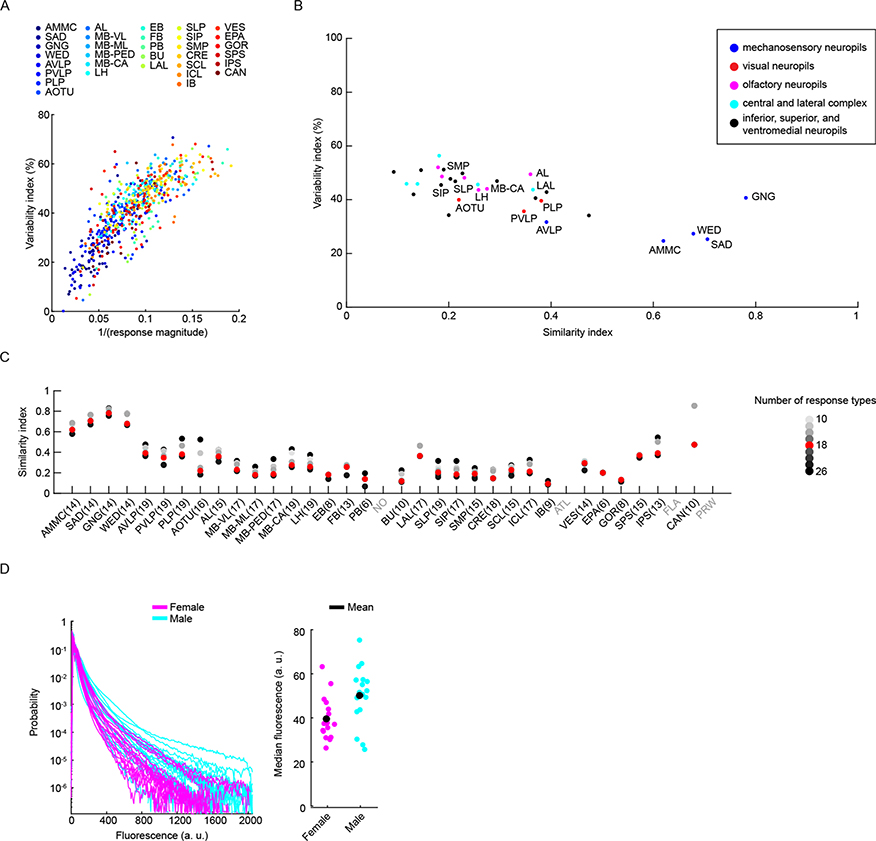
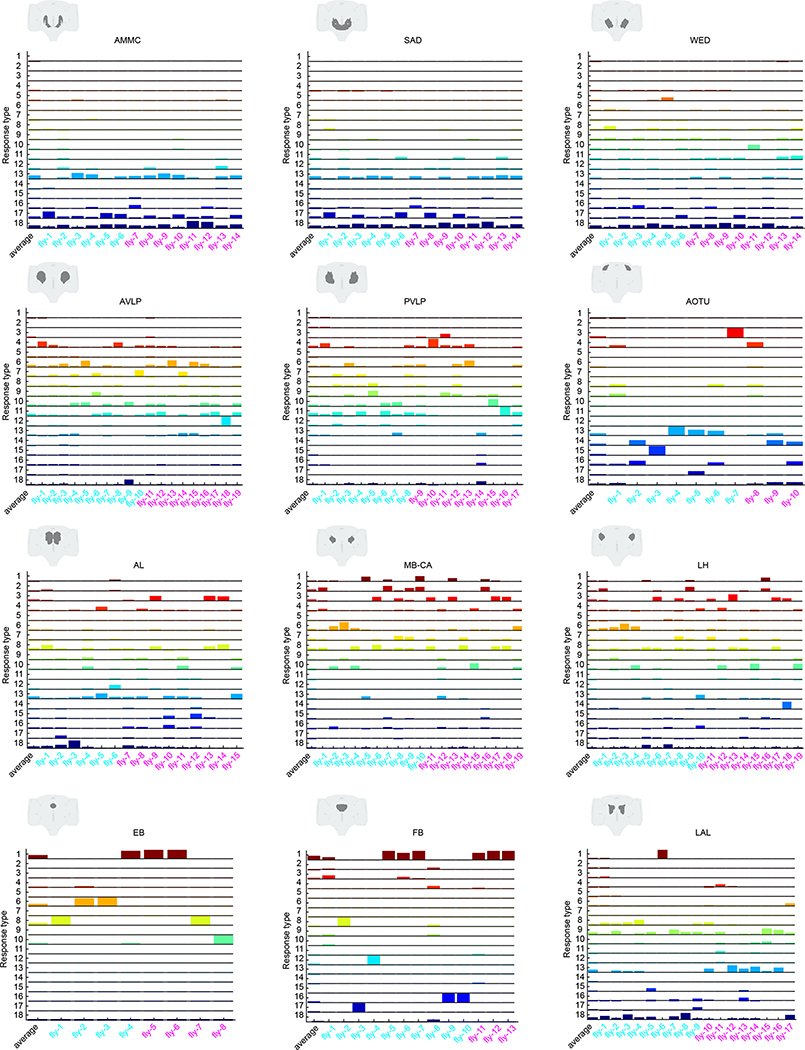
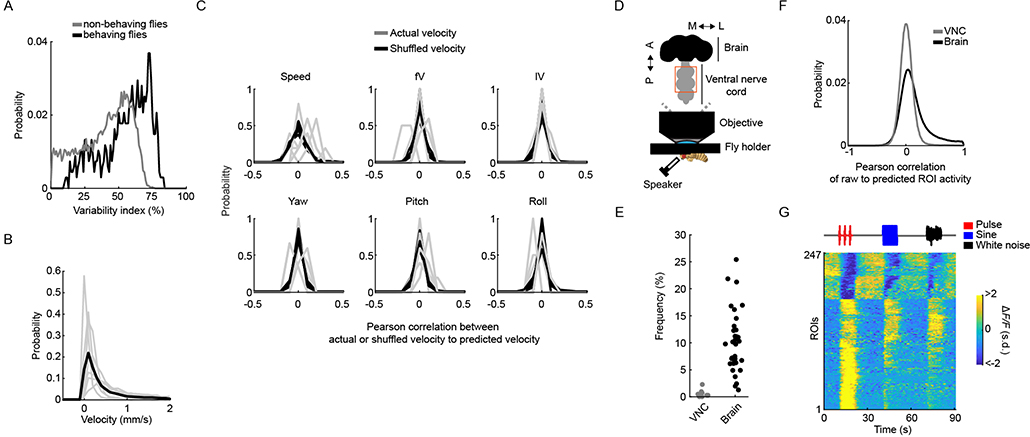
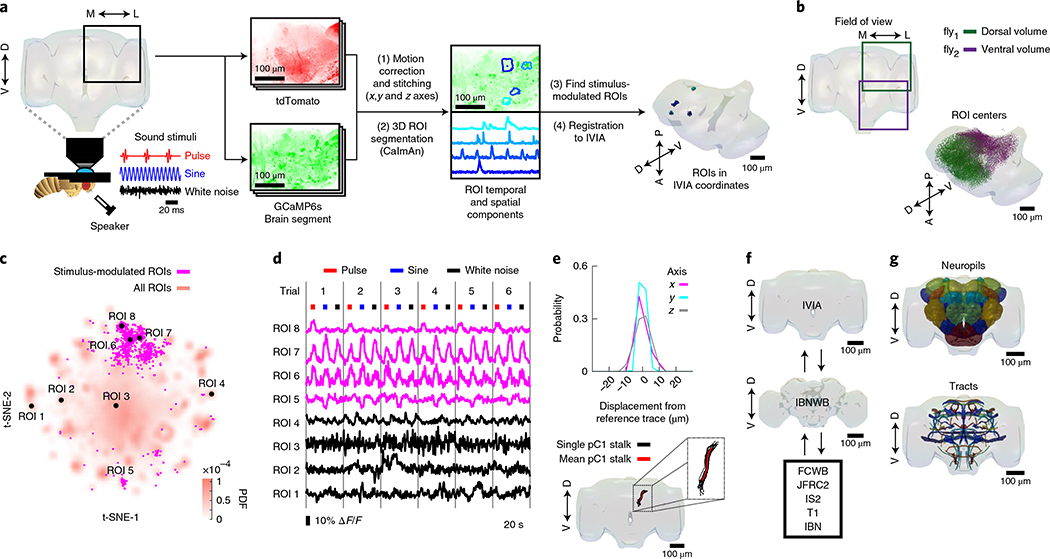
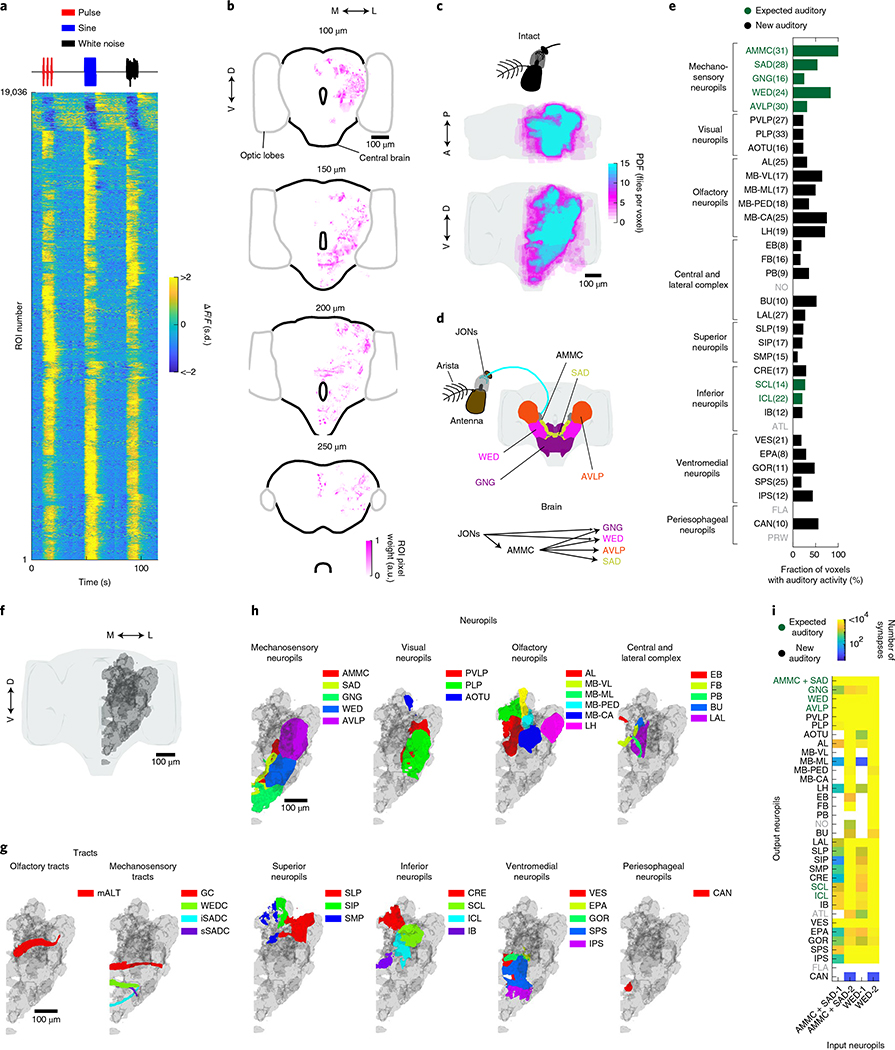
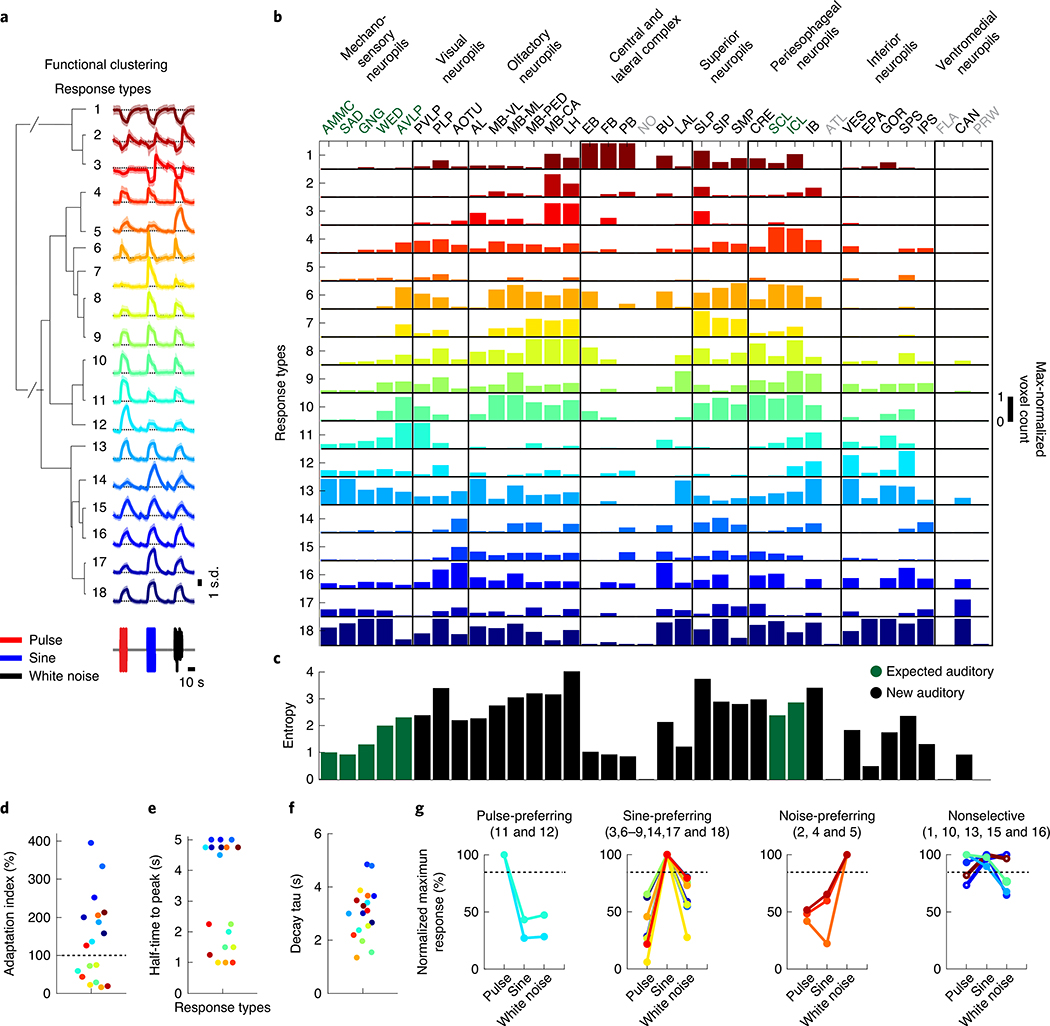
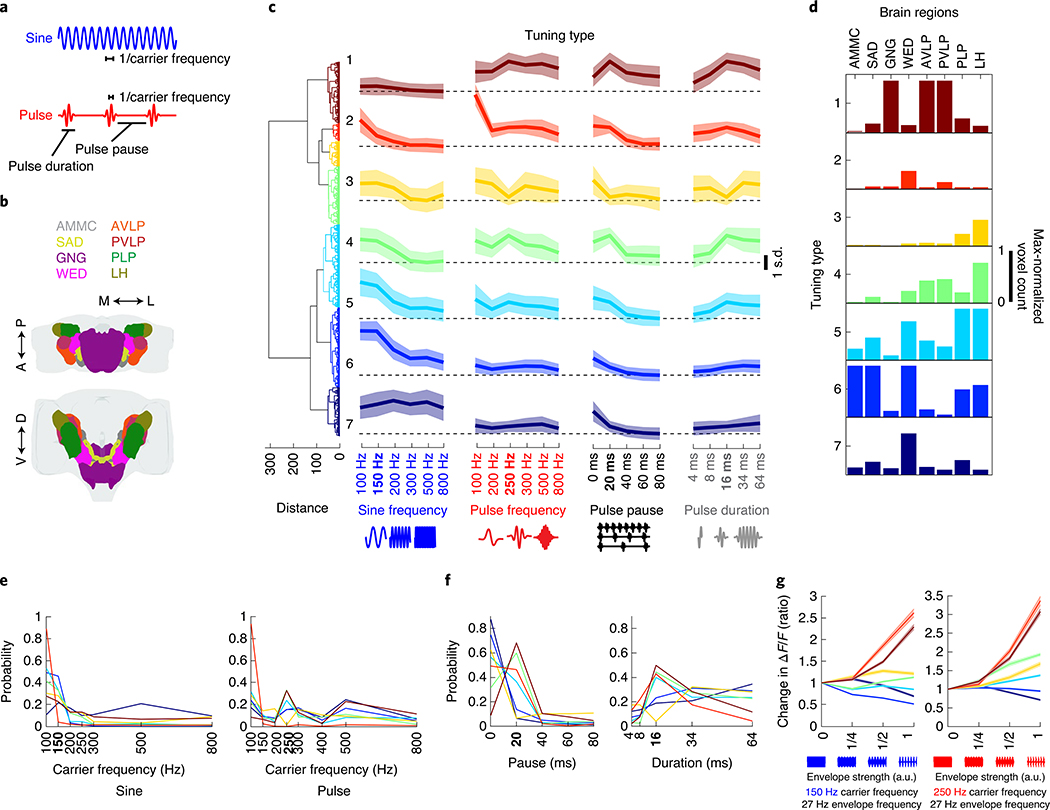
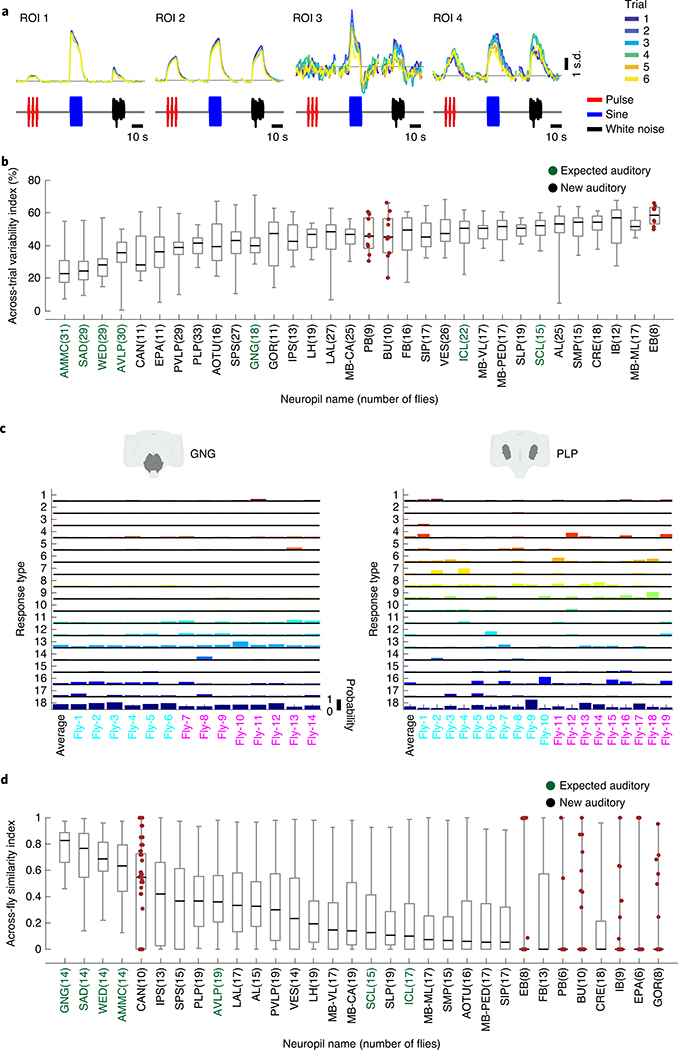
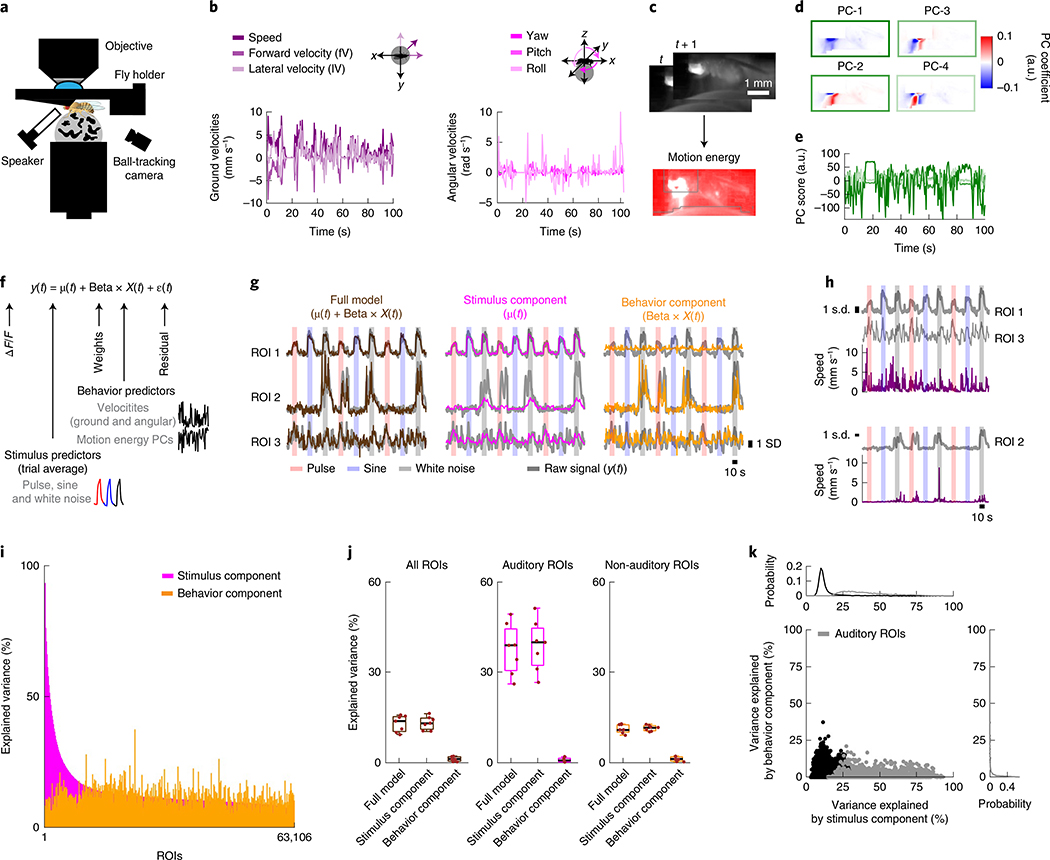
Sensory pathways are typically studied by starting at receptor neurons and following postsynaptic neurons into the brain. However, this leads to a bias in analyses of activity toward the earliest layers of processing. Here, we present new methods for volumetric neural imaging with precise across-brain registration to characterize auditory activity throughout the entire central brain of Drosophila and make comparisons across trials, individuals and sexes. We discover that auditory activity is present in most central brain regions and in neurons responsive to other modalities. Auditory responses are temporally diverse, but the majority of activity is tuned to courtship song features. Auditory responses are stereotyped across trials and animals in early mechanosensory regions, becoming more variable at higher layers of the putative pathway, and this variability is largely independent of ongoing movements. This study highlights the power of using an unbiased, brain-wide approach for mapping the functional organization of sensory activity.
Figures


References
-
- von Schilcher F The role of auditory stimuli in the courtship of Drosophila melanogaster. Anim. Behav 24, 18–26 (1976).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
