

Sex differences and effects of the estrous stage on hippocampal-prefrontal theta communications
- PMID: 33230976
- PMCID: PMC7683809
- DOI: 10.14814/phy2.14646
Sex differences and effects of the estrous stage on hippocampal-prefrontal theta communications
Abstract
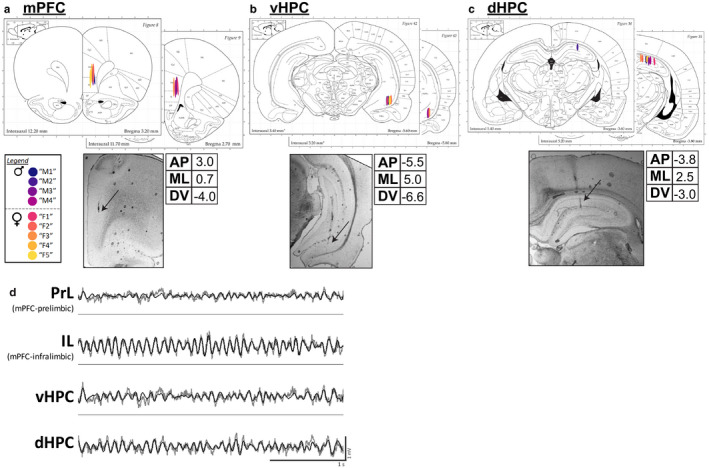
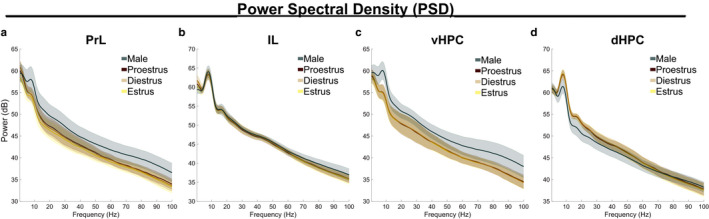
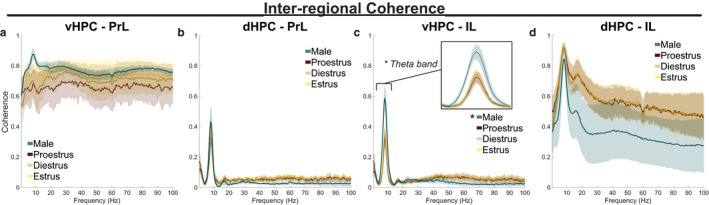
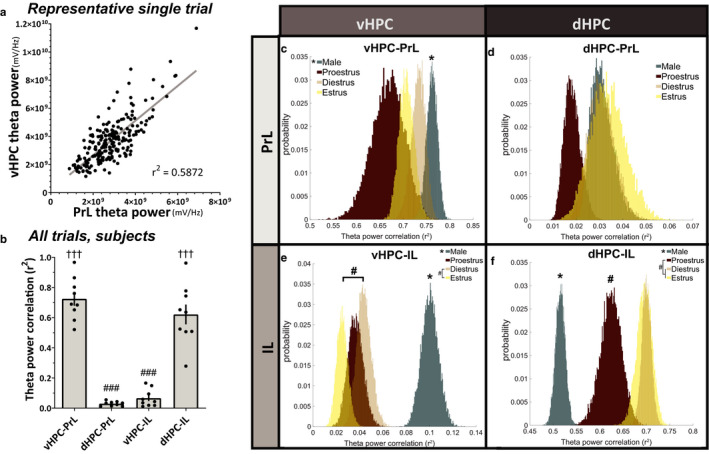
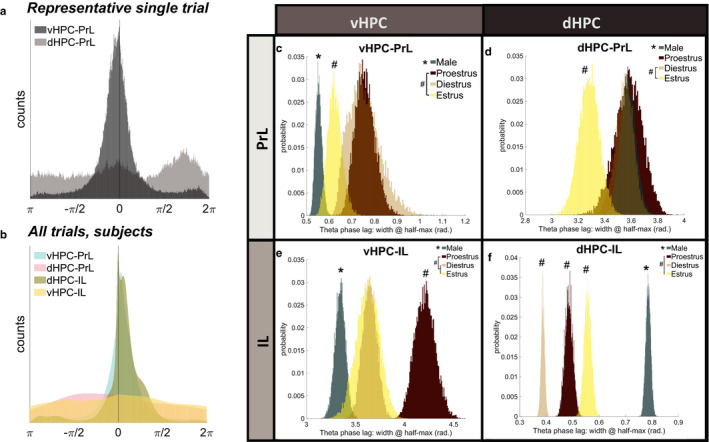
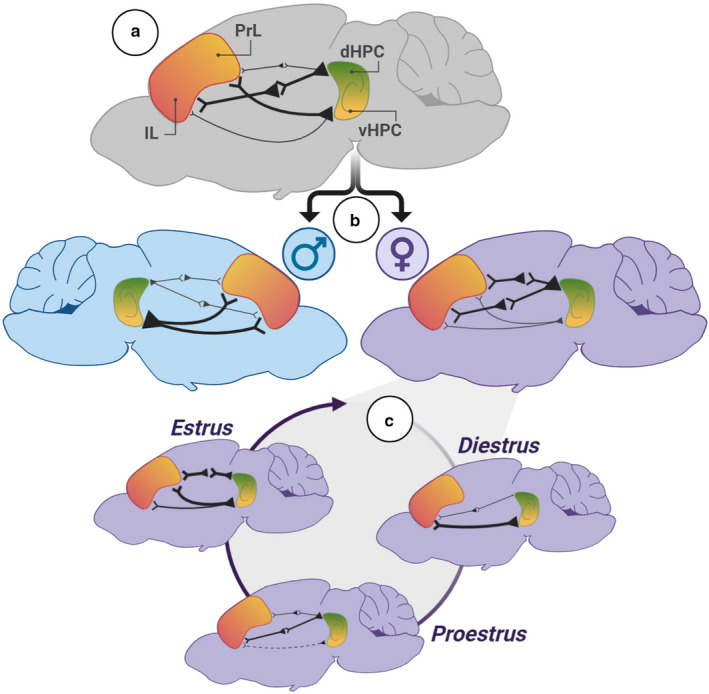
Effective communication between the mammalian hippocampus and neocortex is essential to certain cognitive-behavioral tasks critical to survival in a changing environment. Notably, functional synchrony between local field potentials (LFPs) of the ventral hippocampus (vHPC) and the medial prefrontal cortex (mPFC) within the theta band (4-12 Hz) underlies innate avoidance behavior during approach-avoidance conflict tasks in male rodents. However, the physiology of vHPC-mPFC communications in females remains unestablished. Furthermore, little is known about how mPFC subdivisions functionally interact in the theta band with hippocampal subdivisions in both sexes in the absence of task demand. Given the established roles of biological sex and gonadal hormone status on innate avoidance behaviors and neuronal excitability, here, we characterize the effects of biological sex and female estrous stage on hippocampal-prefrontal (HPC-mPFC) theta signaling in freely moving female and male rats. LFPs from vHPC, dorsal hippocampus (dHPC), mPFC-prelimbic (PrL), and mPFC-infralimbic (IL) were simultaneously recorded during spontaneous exploration of a familiar arena. Data suggest that theta phase and power in vHPC preferentially synchronize with PrL; conversely, dHPC and IL preferentially synchronize. Males displayed greater vHPC-PrL theta synchrony than females, despite similar regional frequency band power and inter-regional coherence. Additionally, several significant estrous-linked changes in HPC-mPFC theta dynamics were observed. These findings support the hypothesis that HPC-mPFC theta signaling is sensitive to both biological sex and female estrous stage. These findings establish novel research avenues concerning sex as a biological variable and effects of gonadal hormone status on HPC-mPFC network activity as it pertains to threat evaluation biomarkers.
Keywords: estrous; sex differences; theta oscillations.
© 2020 The Authors. Physiological Reports published by Wiley Periodicals LLC on behalf of The Physiological Society and the American Physiological Society.
Conflict of interest statement
Authors declare no conflicts of interest.
Figures
References
-
- Becker, J. B. , Arnold, A. P. , Berkley, K. J. , Blaustein, J. D. , Eckel, L. A. , Hampson, E. , Herman, J. P. , Marts, S. , Sadee, W. , Steiner, M. , Taylor, J. , & Young, E. (2005). Strategies and methods for research on sex differences in brain and behavior. Endocrinology, 146, 1650–1673. 10.1210/en.2004-1142 - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
