

Odorant Receptors for Detecting Flowering Plant Cues Are Functionally Conserved across Moths and Butterflies
- PMID: 33231630
- PMCID: PMC8042770
- DOI: 10.1093/molbev/msaa300
Odorant Receptors for Detecting Flowering Plant Cues Are Functionally Conserved across Moths and Butterflies
Abstract
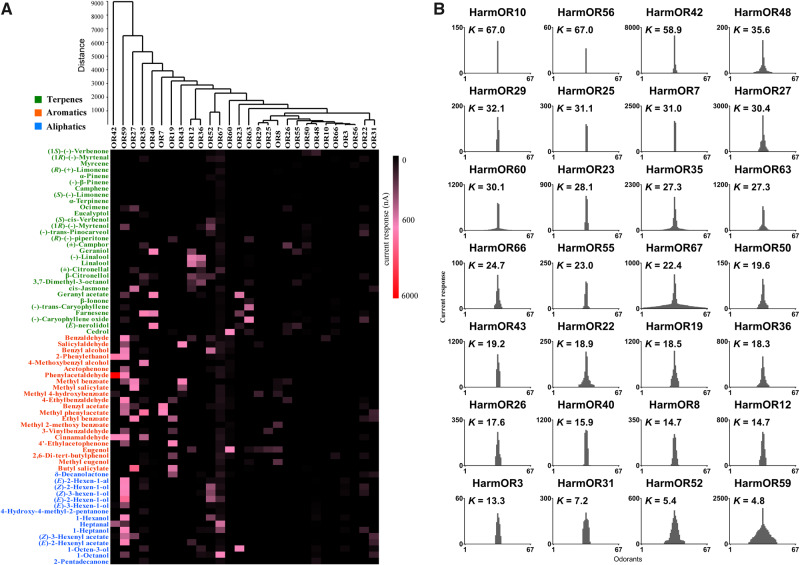
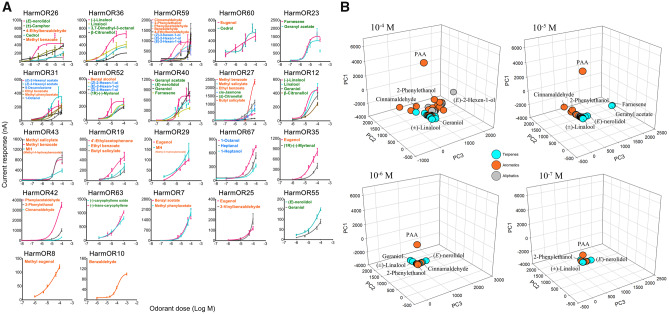
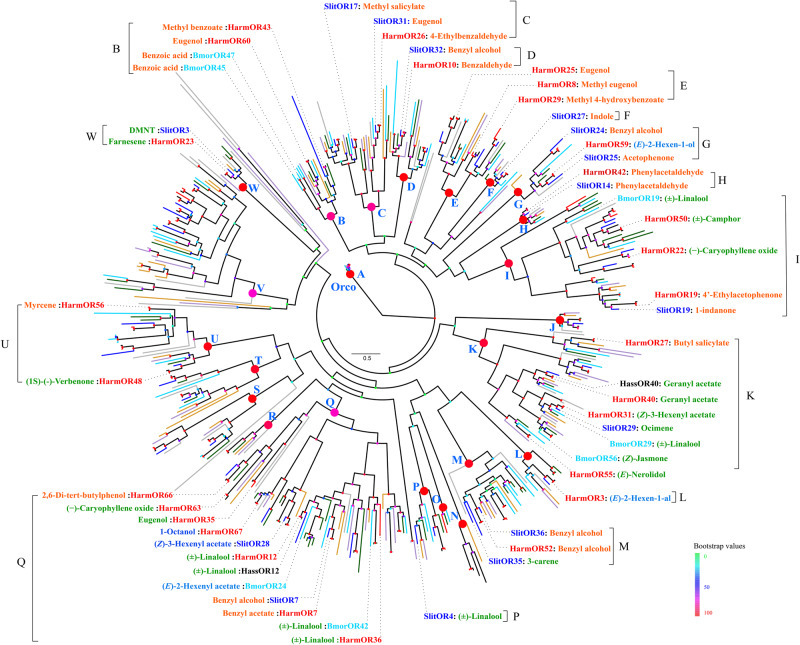
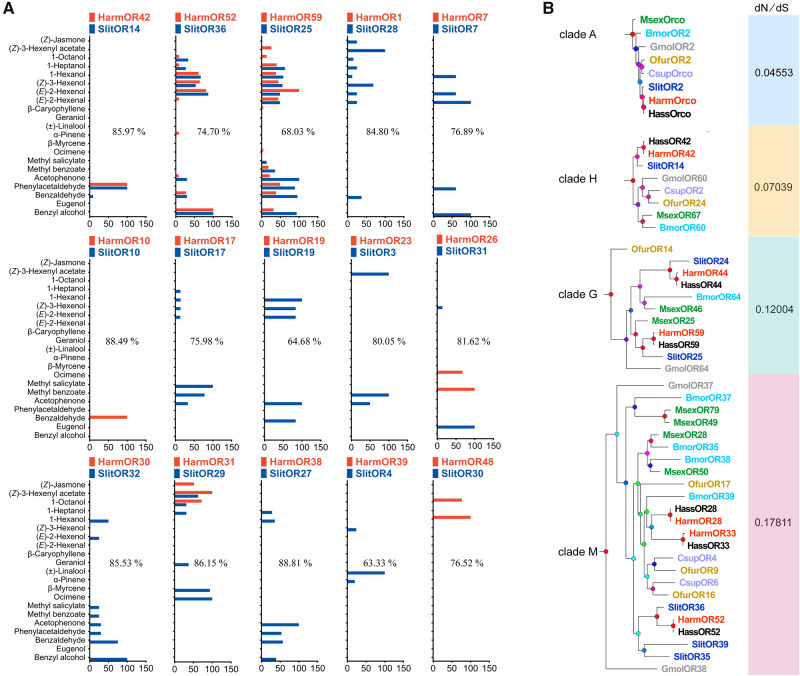
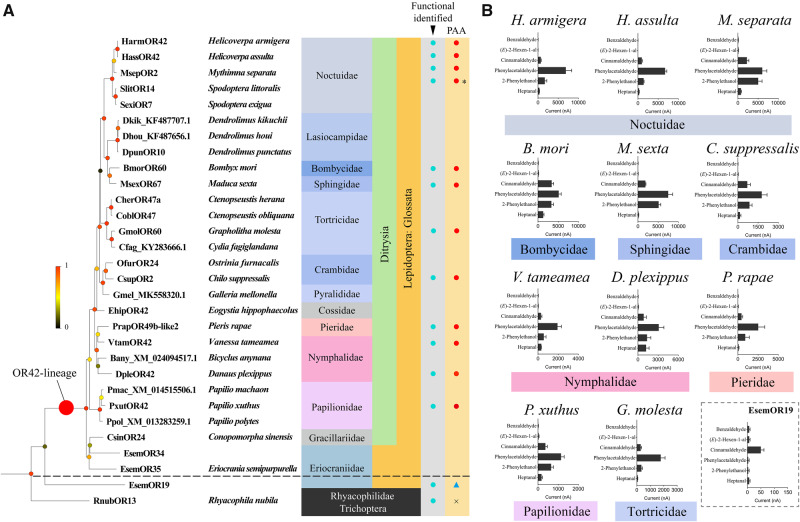
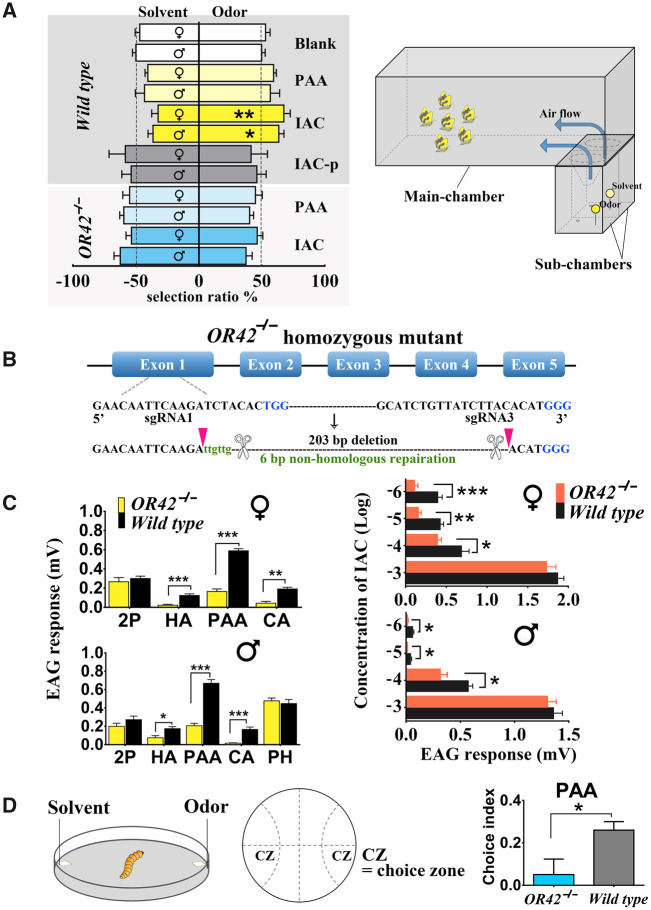
Odorant receptors (ORs) are essential for plant-insect interactions. However, despite the global impacts of Lepidoptera (moths and butterflies) as major herbivores and pollinators, little functional data are available about Lepidoptera ORs involved in plant-volatile detection. Here, we initially characterized the plant-volatile-sensing function(s) of 44 ORs from the cotton bollworm Helicoverpa armigera, and subsequently conducted a large-scale comparative analysis that establishes how most orthologous ORs have functionally diverged among closely related species whereas some rare ORs are functionally conserved. Specifically, our systematic analysis of H. armigera ORs cataloged the wide functional scope of the H. armigera OR repertoire, and also showed that HarmOR42 and its Spodoptera littoralis ortholog are functionally conserved. Pursuing this, we characterized the HarmOR42-orthologous ORs from 11 species across the Glossata suborder and confirmed the HarmOR42 orthologs form a unique OR lineage that has undergone strong purifying selection in Glossata species and whose members are tuned with strong specificity to phenylacetaldehyde, a floral scent component common to most angiosperms. In vivo studies via HarmOR42 knockout support that HarmOR42-related ORs are essential for host-detection by sensing phenylacetaldehyde. Our work also supports that these ORs coevolved with the tube-like proboscis, and has maintained functional stability throughout the long-term coexistence of Lepidoptera with angiosperms. Thus, beyond providing a rich empirical resource for delineating the precise functions of H. armigera ORs, our results enable a comparative analysis of insect ORs that have apparently facilitated and currently sustain the intimate adaptations and ecological interactions among nectar feeding insects and flowering plants.
Keywords: Helicoverpa armigera; Glossata; odorant receptor; phenylacetaldehyde; plant volatile.
© The Author(s) 2020. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Anderson AR, Wanner KW, Trowell SC, Warr CG, Jaquin-Joly E, Zagatti P, Robertson H, Newcomb RD.. 2009. Molecular basis of female-specific odorant responses in Bombyx mori. Insect Biochem Mol Biol. 39(3):189–197. - PubMed
-
- Anderson P, Hilker M, Hansson BS, Bombosch S, Klein B, Schildknecht H.. 1993. Oviposition deterring components in larval frass of Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae): a behavioural and electrophysiological evaluation. J Insect Physiol. 39(2):129–137.
-
- Bruce TJ, Wadhams LJ, Woodcock CM.. 2005. Insect host location: a volatile situation. Trends Plant Sci. 10(6):269–274. - PubMed
