

Sleep Promotes Downward Firing Rate Homeostasis
- PMID: 33232655
- PMCID: PMC7864886
- DOI: 10.1016/j.neuron.2020.11.001
Sleep Promotes Downward Firing Rate Homeostasis
Abstract
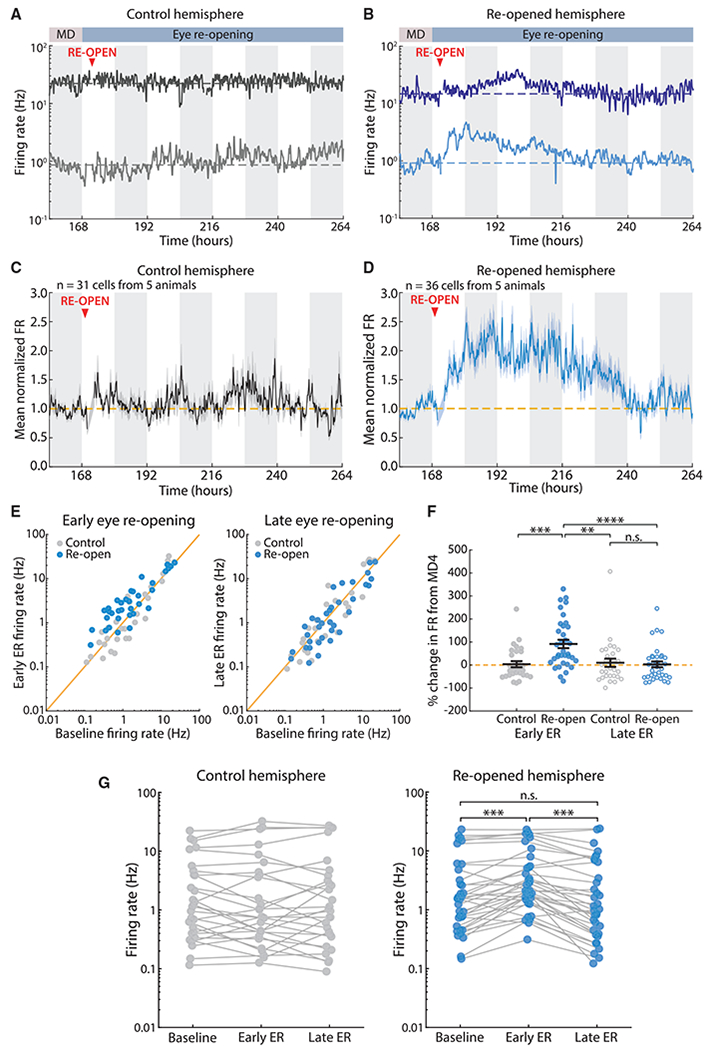
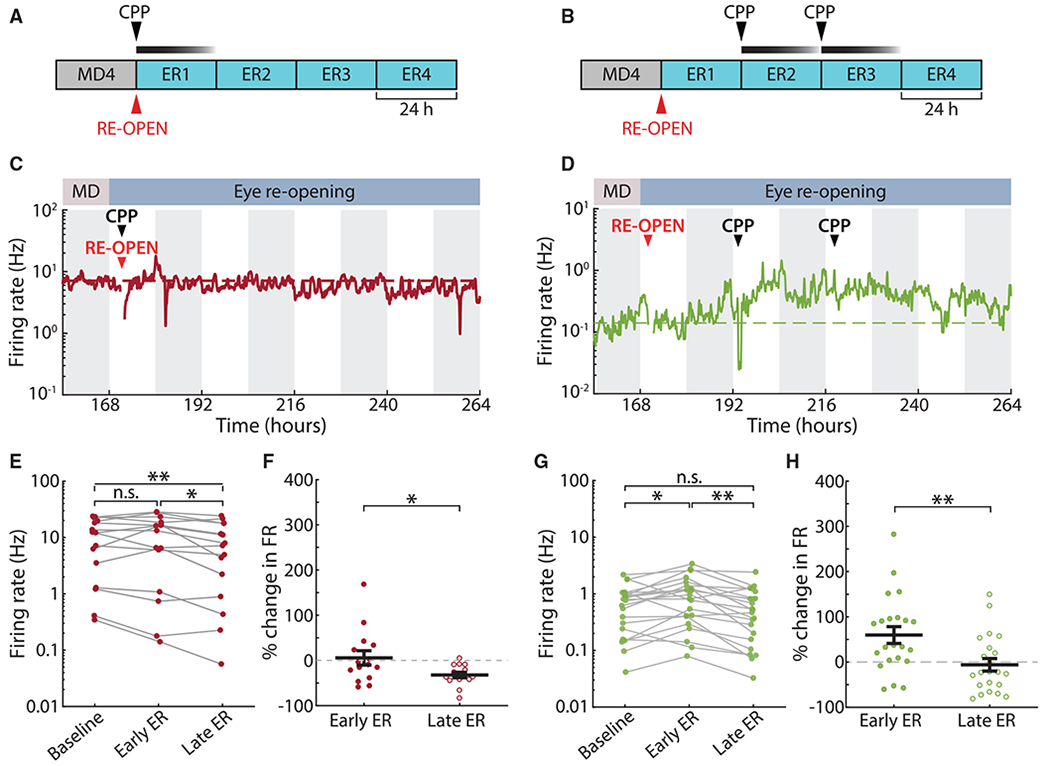
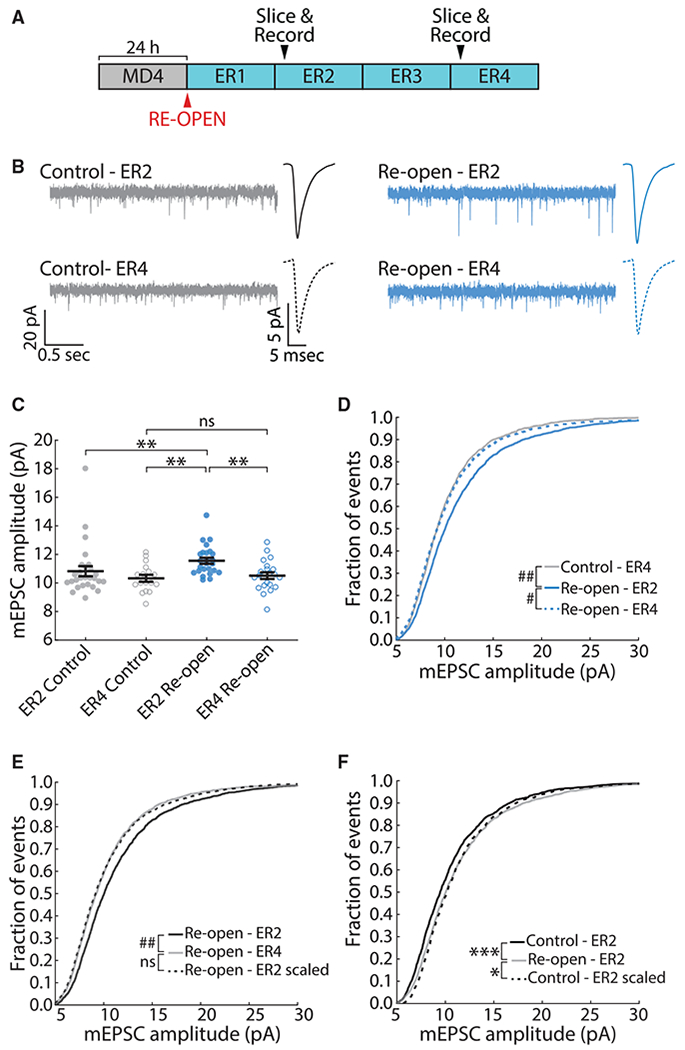
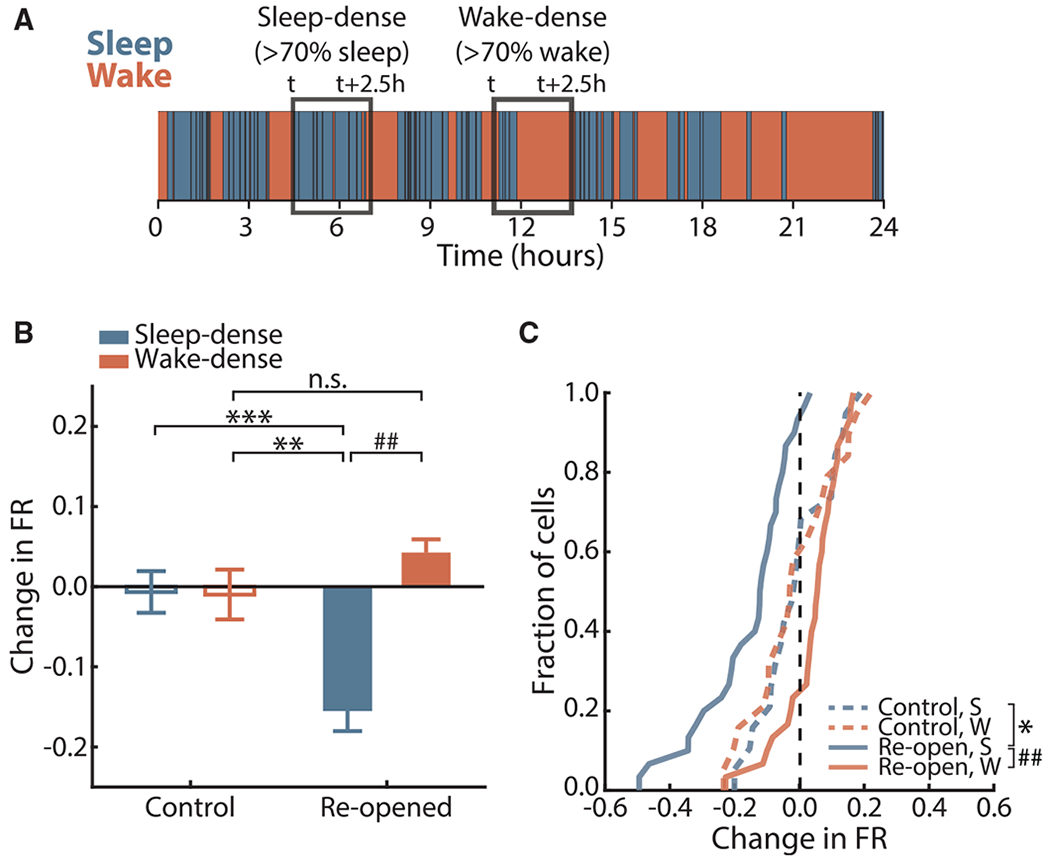
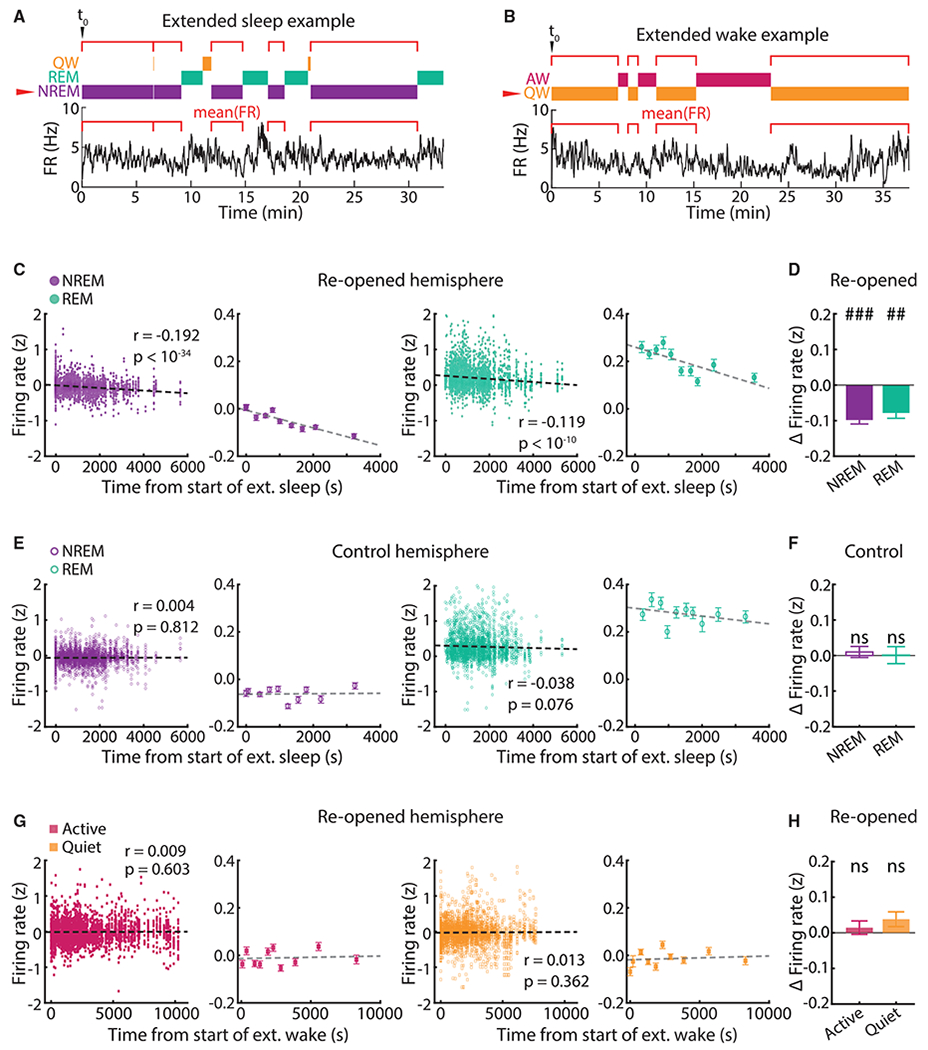
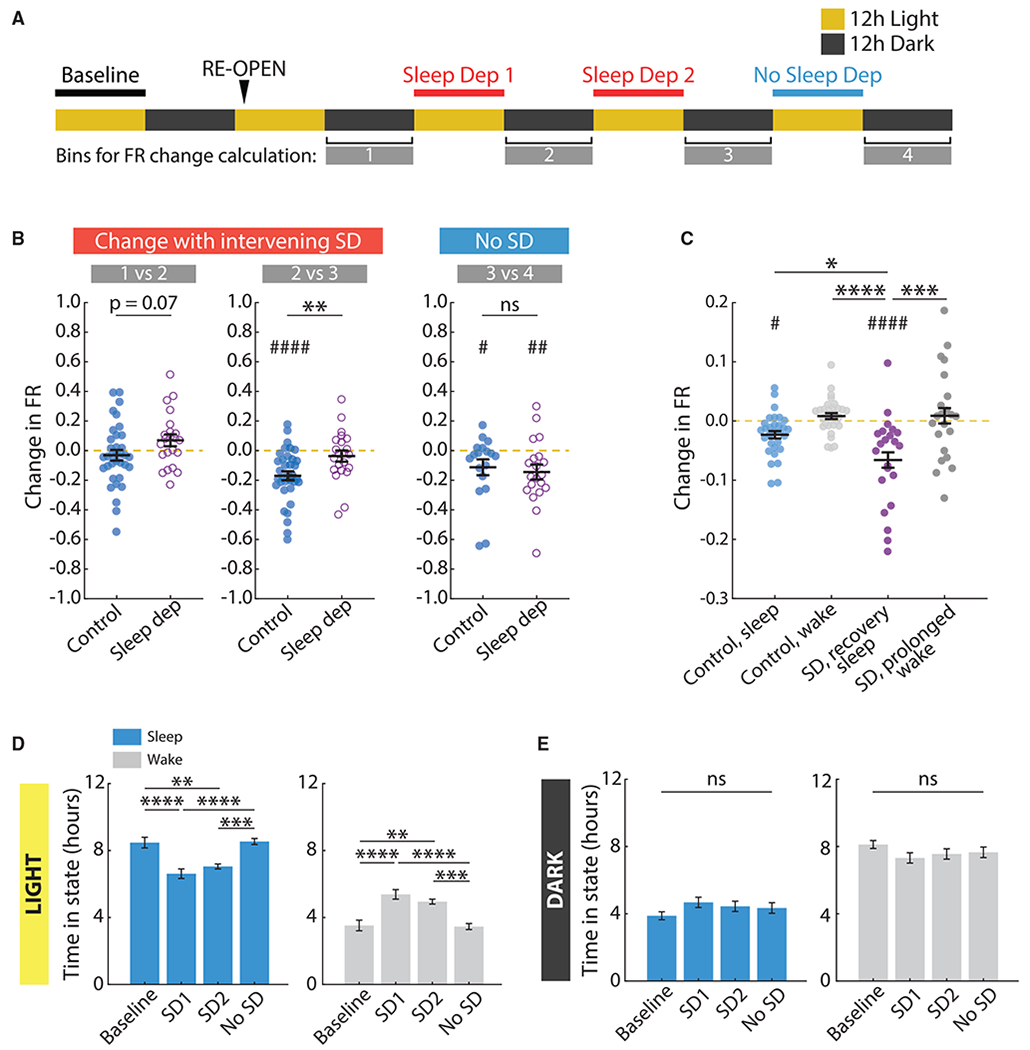
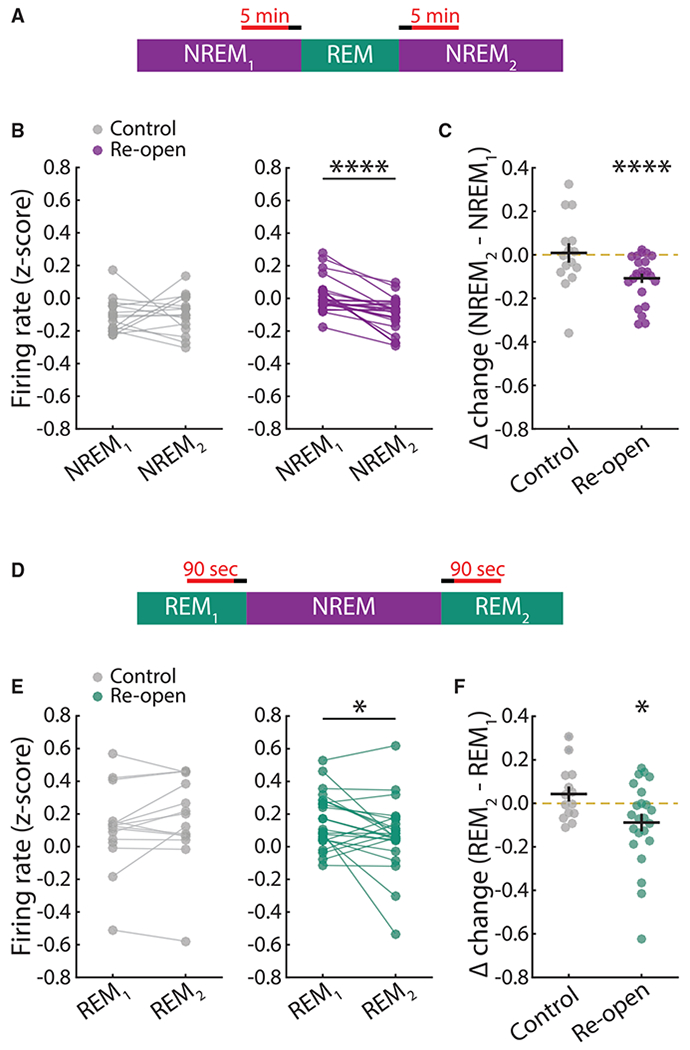
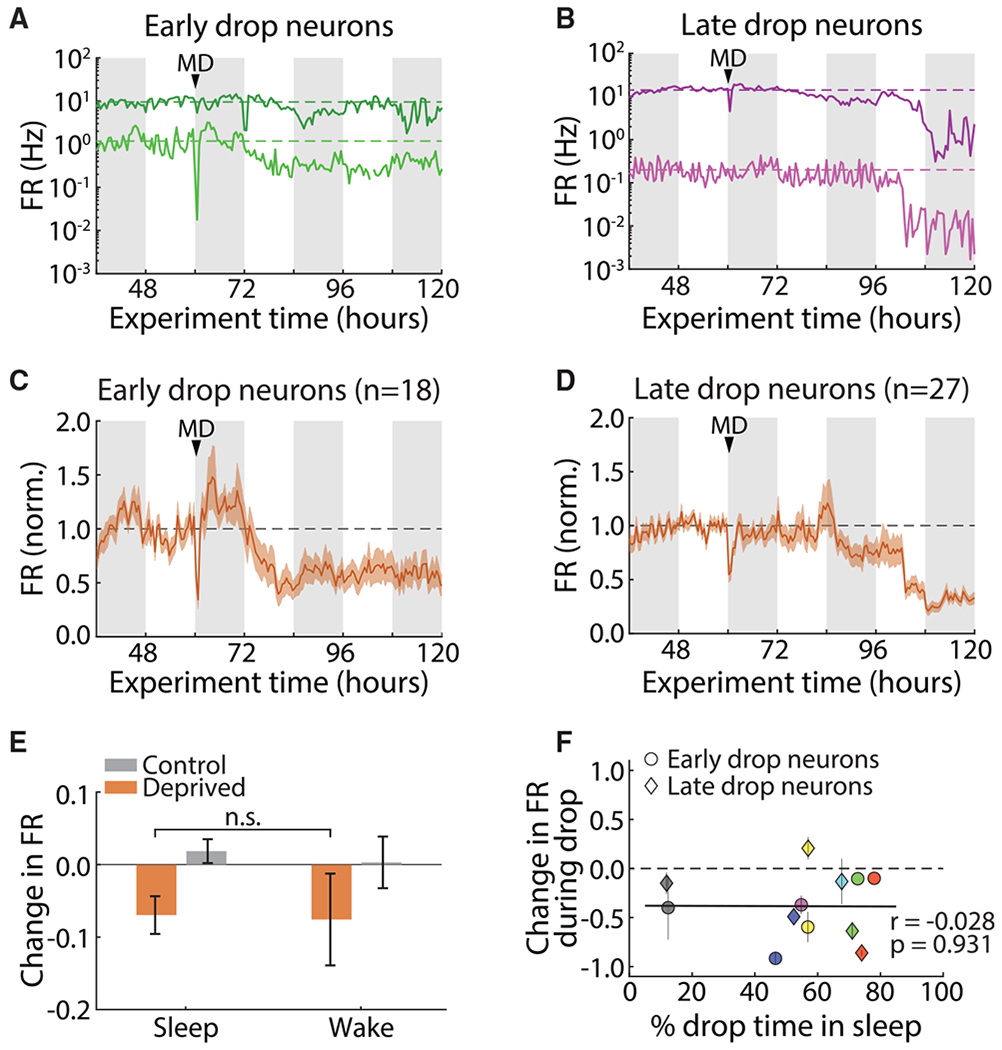
Homeostatic plasticity is hypothesized to bidirectionally regulate neuronal activity around a stable set point to compensate for learning-related plasticity, but to date only upward firing rate homeostasis (FRH) has been demonstrated in vivo. We combined chronic electrophysiology in freely behaving animals with an eye-reopening paradigm to enhance firing in primary visual cortex (V1) and found that neurons bidirectionally regulate firing rates around an individual set point. Downward FRH did not require N-methyl-D-aspartate receptor (NMDAR) signaling and was associated with homeostatic scaling down of synaptic strengths. Like upward FRH, downward FRH was gated by arousal state but in the opposite direction: it occurred during sleep, not during wake. In contrast, firing rate depression associated with Hebbian plasticity happened independently of sleep and wake. Thus, sleep and wake states temporally segregate upward and downward FRH, which might prevent interference or provide unopposed homeostatic compensation when it is needed most.
Keywords: Experience-dependent plasticity; Firing rate homeostasis; Homeostatic plasticity; Sleep; Synaptic scaling; Visual system.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests G.G.T. is a member of the Neuron advisory board. The authors declare no competing interests.
Figures
Comment in
-
Sleep calms firing rates.Nat Rev Neurosci. 2021 Feb;22(2):74-75. doi: 10.1038/s41583-020-00419-3. Nat Rev Neurosci. 2021. PMID: 33293691 No abstract available.
-
The Ups and Downs of Firing Rate Homeostasis.Neuron. 2021 Feb 3;109(3):401-403. doi: 10.1016/j.neuron.2020.12.024. Neuron. 2021. PMID: 33539775
References
-
- Abbott LF, and Nelson SB (2000). Synaptic plasticity: taming the beast. Nat. Neurosci 3 (Suppl), 1178–1183. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
