

Interplay between Hepatitis D Virus and the Interferon Response
- PMID: 33233762
- PMCID: PMC7699955
- DOI: 10.3390/v12111334
Interplay between Hepatitis D Virus and the Interferon Response
Abstract
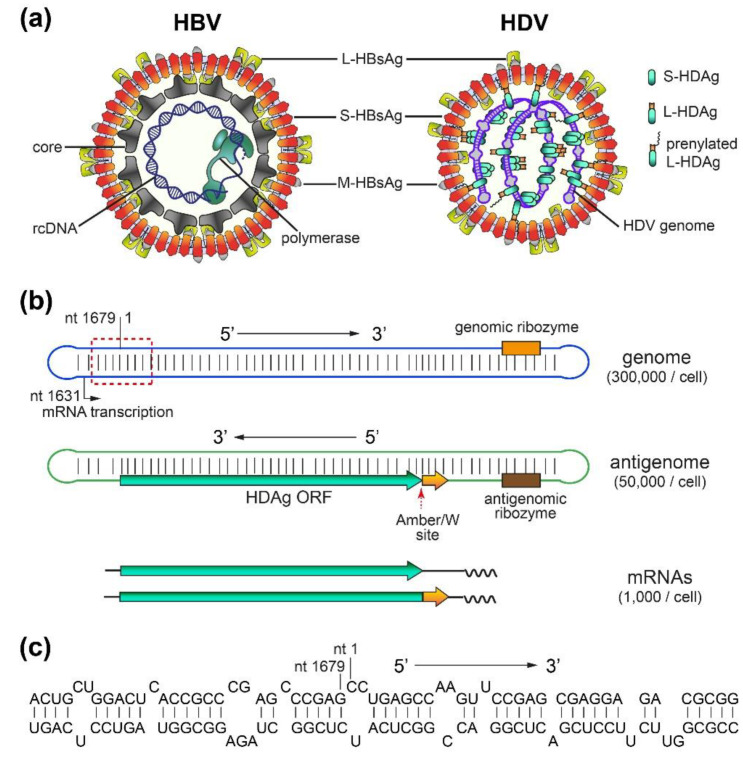
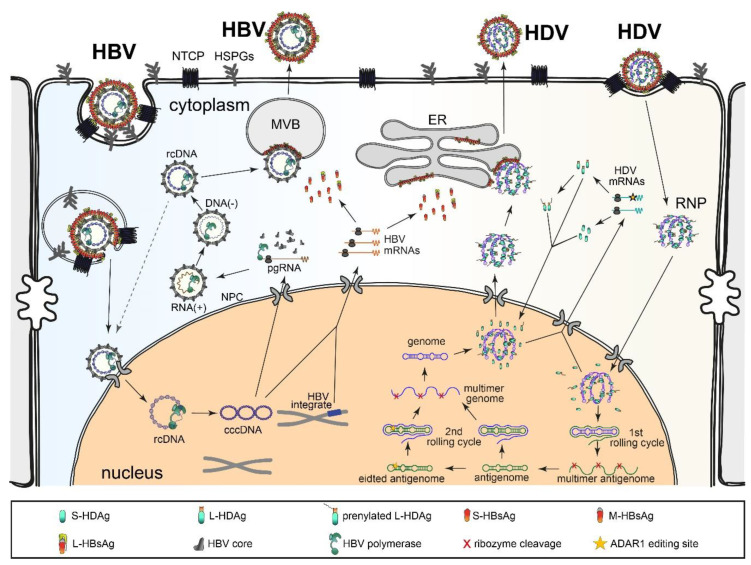
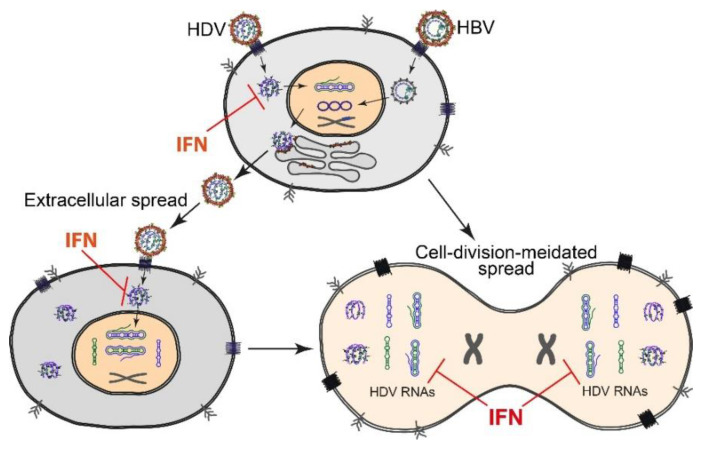
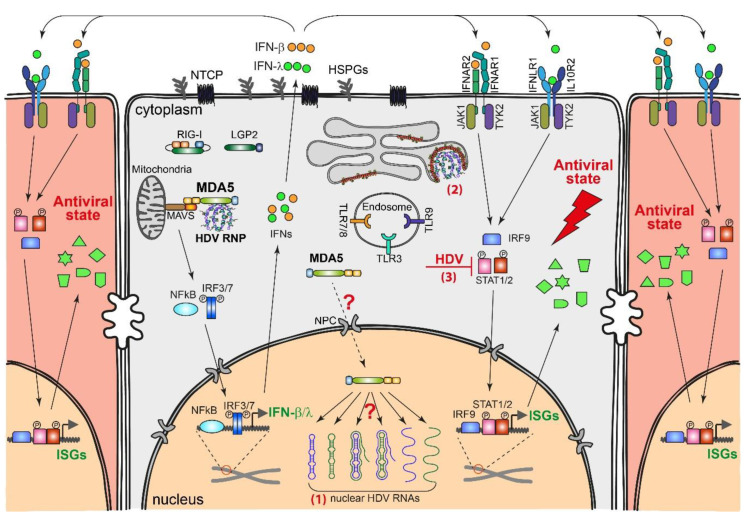
Chronic hepatitis D (CHD) is the most severe form of viral hepatitis, with rapid progression of liver-related diseases and high rates of development of hepatocellular carcinoma. The causative agent, hepatitis D virus (HDV), contains a small (approximately 1.7 kb) highly self-pairing single-strand circular RNA genome that assembles with the HDV antigen to form a ribonucleoprotein (RNP) complex. HDV depends on hepatitis B virus (HBV) envelope proteins for envelopment and de novo hepatocyte entry; however, its intracellular RNA replication is autonomous. In addition, HDV can amplify HBV independently through cell division. Cellular innate immune responses, mainly interferon (IFN) response, are crucial for controlling invading viruses, while viruses counteract these responses to favor their propagation. In contrast to HBV, HDV activates profound IFN response through the melanoma differentiation antigen 5 (MDA5) pathway. This cellular response efficiently suppresses cell-division-mediated HDV spread and, to some extent, early stages of HDV de novo infection, but only marginally impairs RNA replication in resting hepatocytes. In this review, we summarize the current knowledge on HDV structure, replication, and persistence and subsequently focus on the interplay between HDV and IFN response, including IFN activation, sensing, antiviral effects, and viral countermeasures. Finally, we discuss crosstalk with HBV.
Keywords: Hepcludex; Myrcludex B; cell-division-mediated spread; countermeasures; de novo infection; hepatitis B virus; hepatitis D virus; interferon response; pattern recognition receptors; persistence.
Conflict of interest statement
Z.Z. declares no conflicts of interest. S.U. is a co-applicant and co-inventor of patents protecting Hepcludex/bulevirtide. S.U. is a consultant for Gilead Sciences, Myr-GmbH, Galapagos NV, and Humabs BioMed SA. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
