

Potential Rhodopsin- and Bacteriochlorophyll-Based Dual Phototrophy in a High Arctic Glacier
- PMID: 33234687
- PMCID: PMC7701988
- DOI: 10.1128/mBio.02641-20
Potential Rhodopsin- and Bacteriochlorophyll-Based Dual Phototrophy in a High Arctic Glacier
Abstract
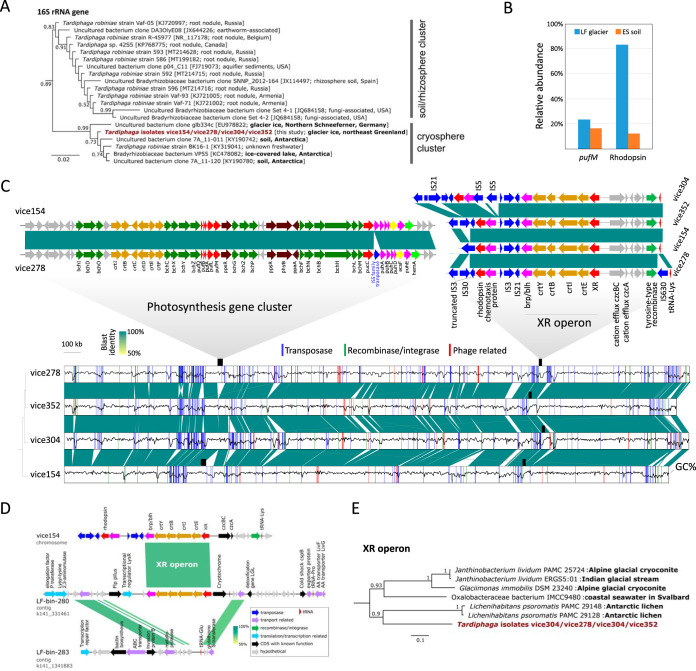
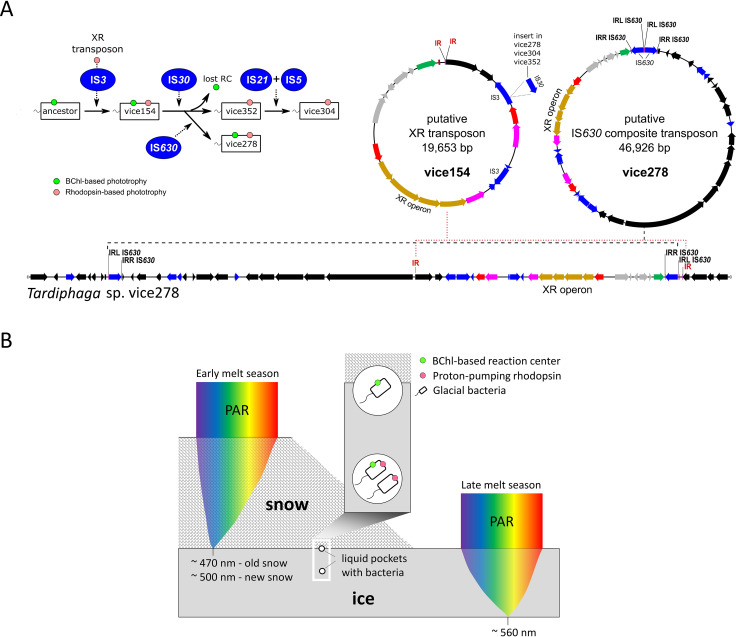
Conserving additional energy from sunlight through bacteriochlorophyll (BChl)-based reaction center or proton-pumping rhodopsin is a highly successful life strategy in environmental bacteria. BChl and rhodopsin-based systems display contrasting characteristics in the size of coding operon, cost of biosynthesis, ease of expression control, and efficiency of energy production. This raises an intriguing question of whether a single bacterium has evolved the ability to perform these two types of phototrophy complementarily according to energy needs and environmental conditions. Here, we report four Tardiphaga sp. strains (Alphaproteobacteria) of monophyletic origin isolated from a high Arctic glacier in northeast Greenland (81.566° N, 16.363° W) that are at different evolutionary stages concerning phototrophy. Their >99.8% identical genomes contain footprints of horizontal operon transfer (HOT) of the complete gene clusters encoding BChl- and xanthorhodopsin (XR)-based dual phototrophy. Two strains possess only a complete XR operon, while the other two strains have both a photosynthesis gene cluster and an XR operon in their genomes. All XR operons are heavily surrounded by mobile genetic elements and are located close to a tRNA gene, strongly signaling that a HOT event of the XR operon has occurred recently. Mining public genome databases and our high Arctic glacial and soil metagenomes revealed that phylogenetically diverse bacteria have the metabolic potential of performing BChl- and rhodopsin-based dual phototrophy. Our data provide new insights on how bacteria cope with the harsh and energy-deficient environment in surface glacier, possibly by maximizing the capability of exploiting solar energy.IMPORTANCE Over the course of evolution for billions of years, bacteria that are capable of light-driven energy production have occupied every corner of surface Earth where sunlight can reach. Only two general biological systems have evolved in bacteria to be capable of net energy conservation via light harvesting: one is based on the pigment of (bacterio-)chlorophyll and the other is based on proton-pumping rhodopsin. There is emerging genomic evidence that these two rather different systems can coexist in a single bacterium to take advantage of their contrasting characteristics in the number of genes involved, biosynthesis cost, ease of expression control, and efficiency of energy production and thus enhance the capability of exploiting solar energy. Our data provide the first clear-cut evidence that such dual phototrophy potentially exists in glacial bacteria. Further public genome mining suggests this understudied dual phototrophic mechanism is possibly more common than our data alone suggested.
Keywords: bacteriochlorophyll; genome evolution; glacial bacteria; phototrophy; rhodopsin.
Copyright © 2020 Zeng et al.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
