

Structure of bacterial phospholipid transporter MlaFEDB with substrate bound
- PMID: 33236984
- PMCID: PMC7790496
- DOI: 10.7554/eLife.62518
Structure of bacterial phospholipid transporter MlaFEDB with substrate bound
Abstract
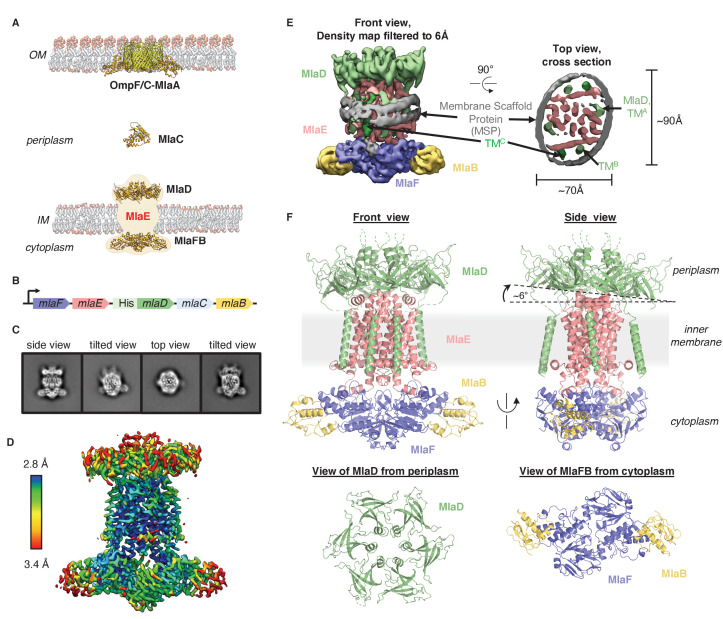
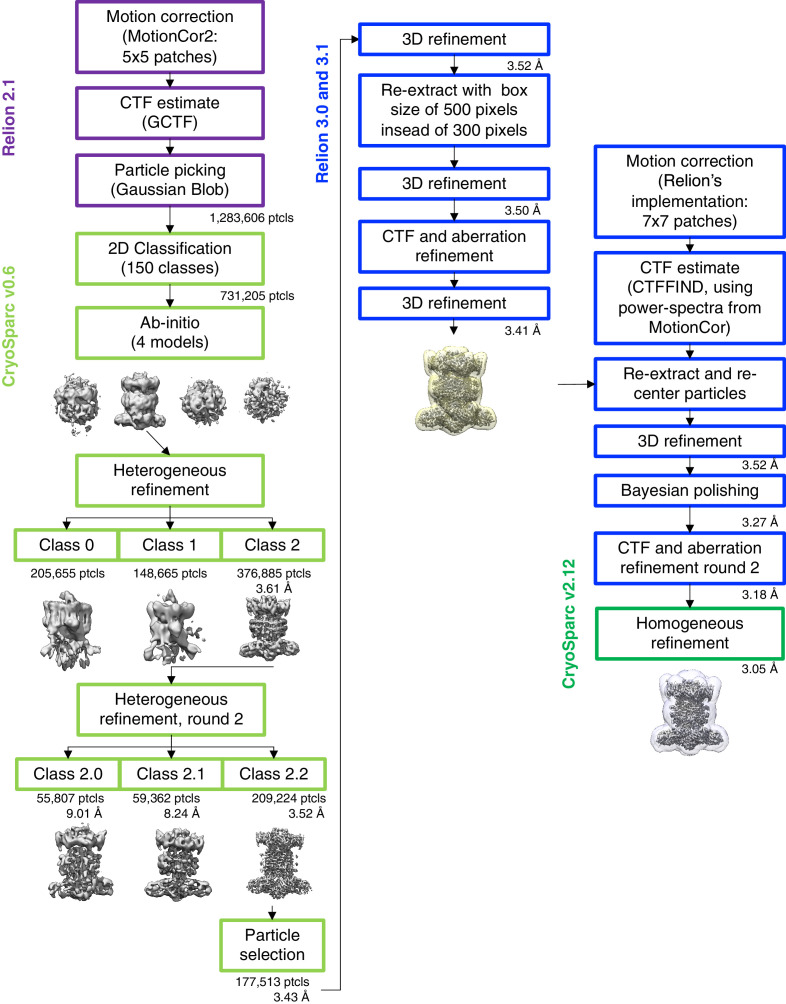
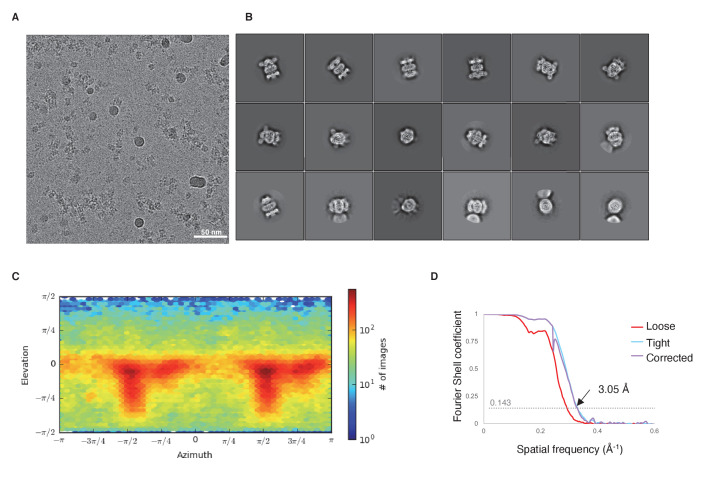
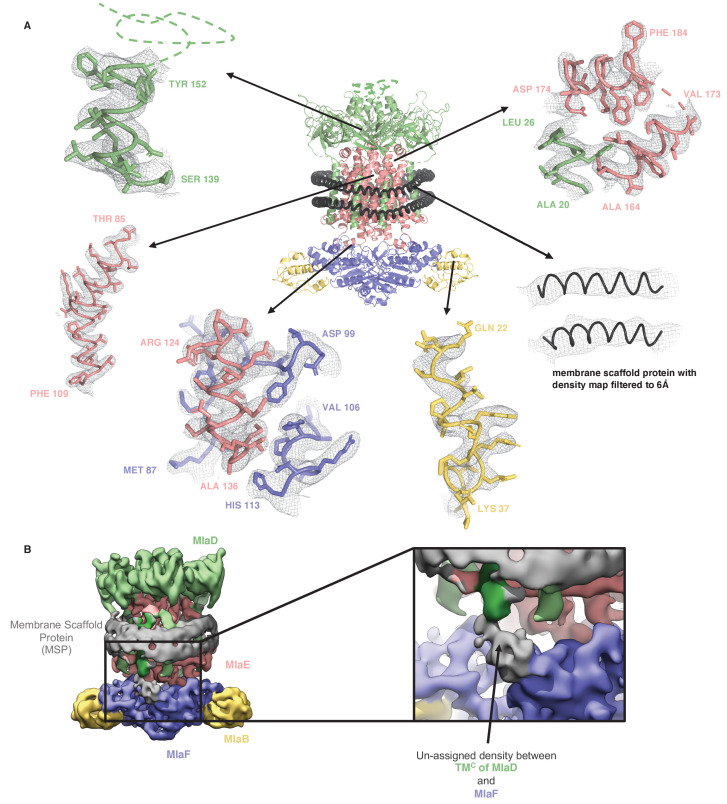
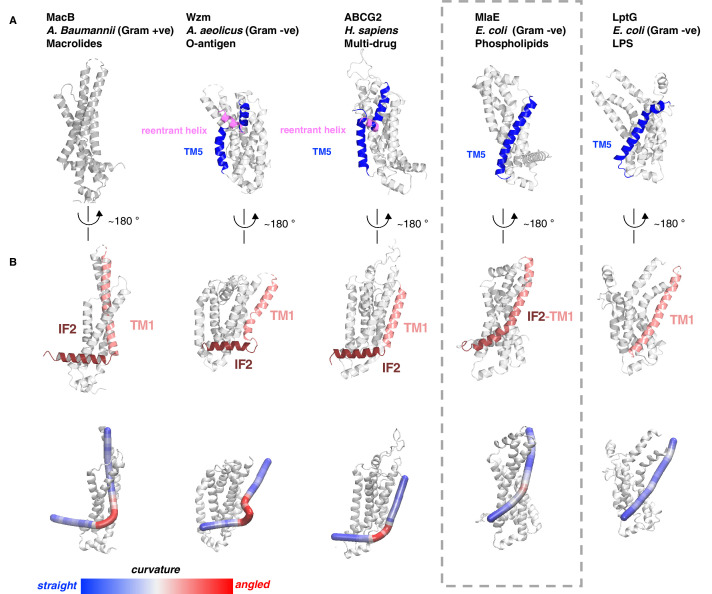
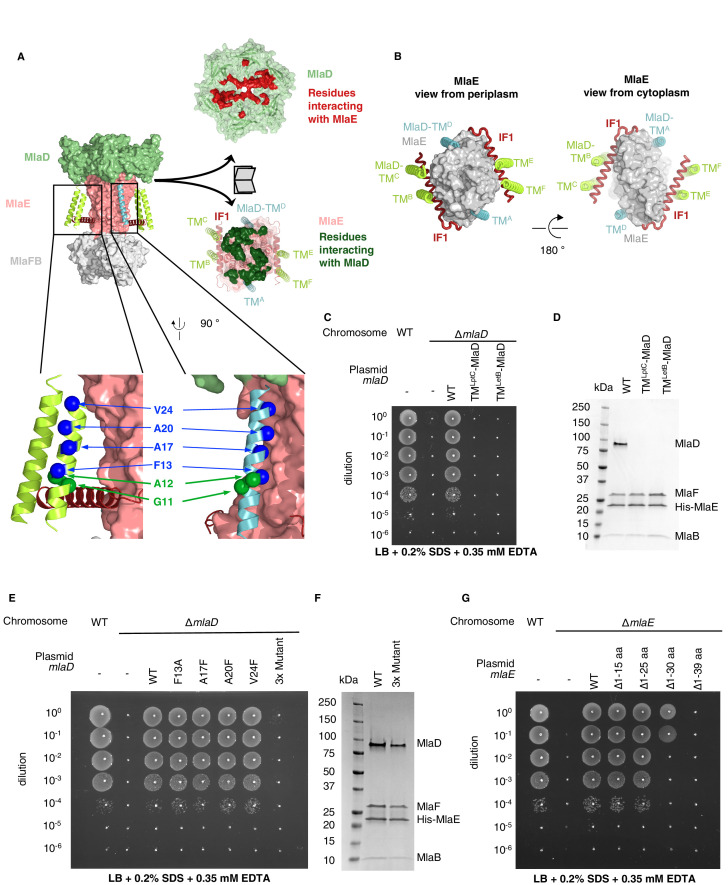
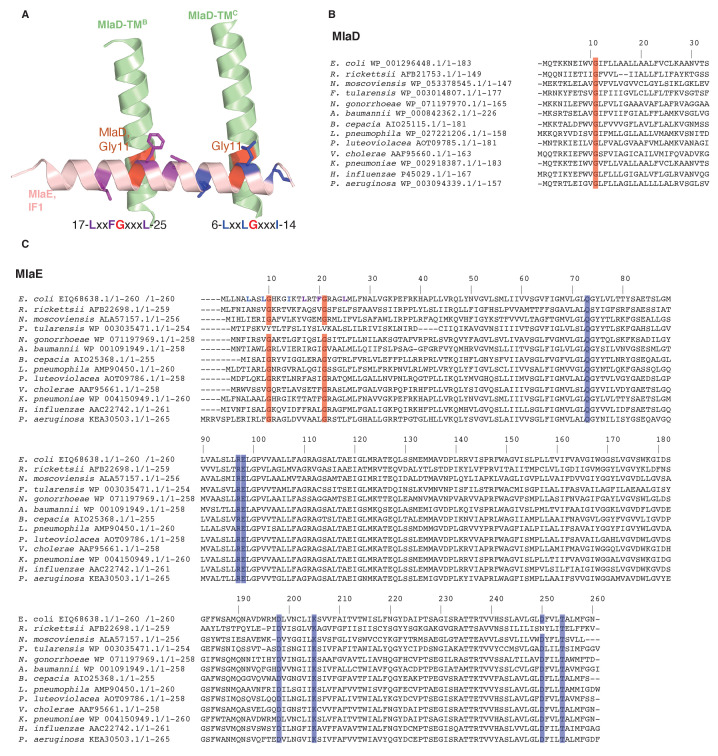
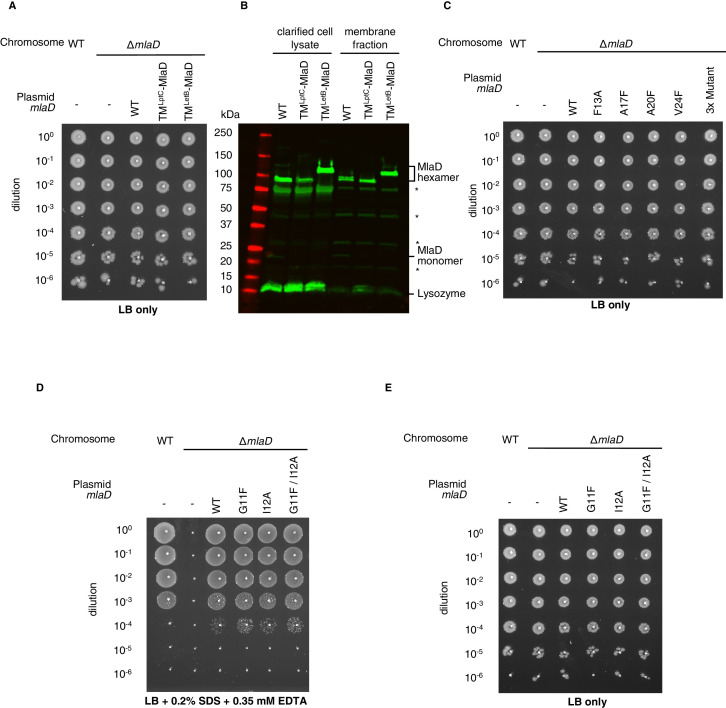
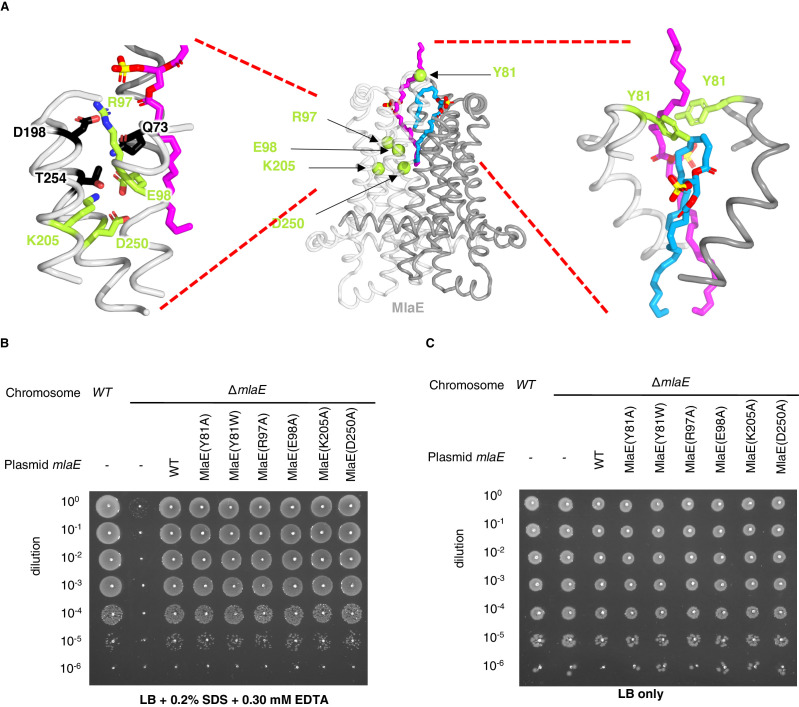
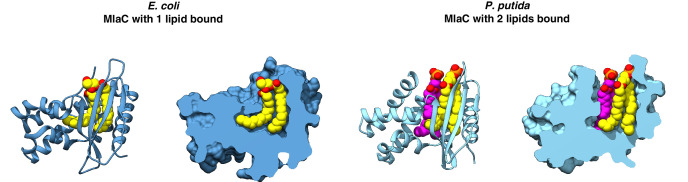
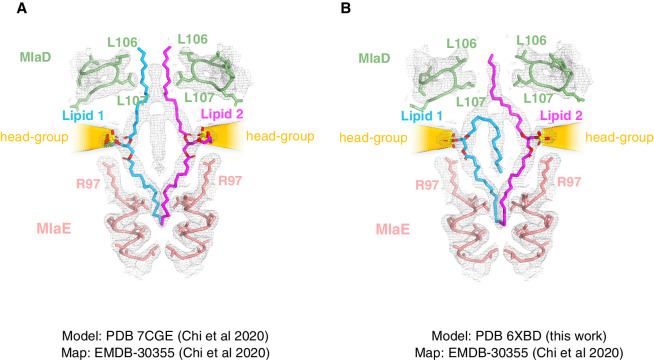
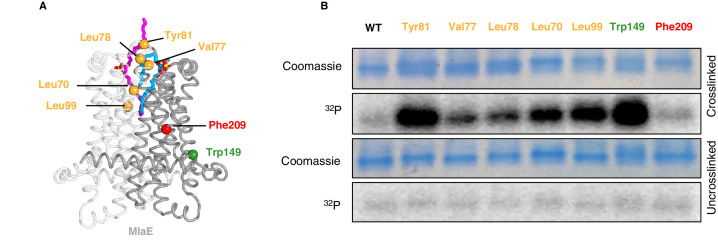
In double-membraned bacteria, phospholipid transport across the cell envelope is critical to maintain the outer membrane barrier, which plays a key role in virulence and antibiotic resistance. An MCE transport system called Mla has been implicated in phospholipid trafficking and outer membrane integrity, and includes an ABC transporter, MlaFEDB. The transmembrane subunit, MlaE, has minimal sequence similarity to other transporters, and the structure of the entire inner-membrane MlaFEDB complex remains unknown. Here, we report the cryo-EM structure of MlaFEDB at 3.05 Å resolution, revealing distant relationships to the LPS and MacAB transporters, as well as the eukaryotic ABCA/ABCG families. A continuous transport pathway extends from the MlaE substrate-binding site, through the channel of MlaD, and into the periplasm. Unexpectedly, two phospholipids are bound to MlaFEDB, suggesting that multiple lipid substrates may be transported each cycle. Our structure provides mechanistic insight into substrate recognition and transport by MlaFEDB.
Keywords: E. coli; bacterial outer membrane; cryo-EM; lipid transport; mla pathway; molecular biophysics; structural biology.
© 2020, Coudray et al.
Conflict of interest statement
NC, GI, MM, MS, GB, DE No competing interests declared
Figures
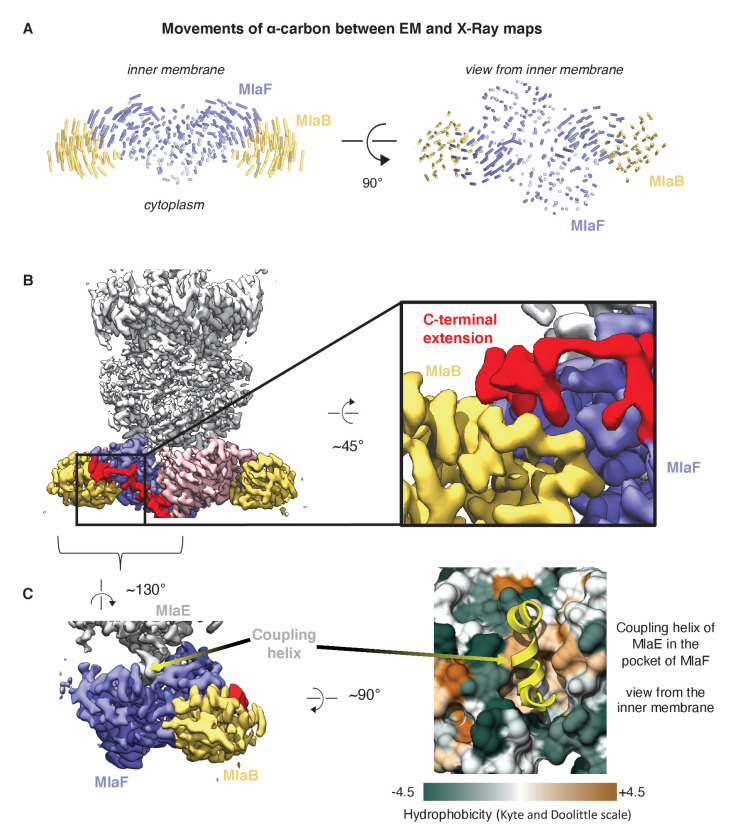
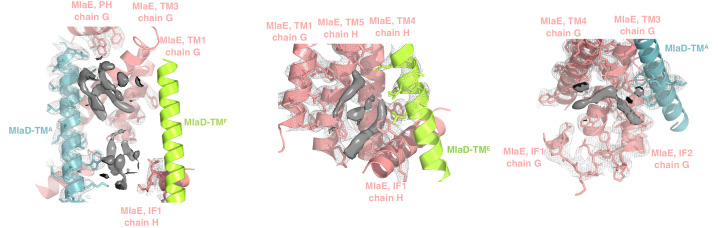
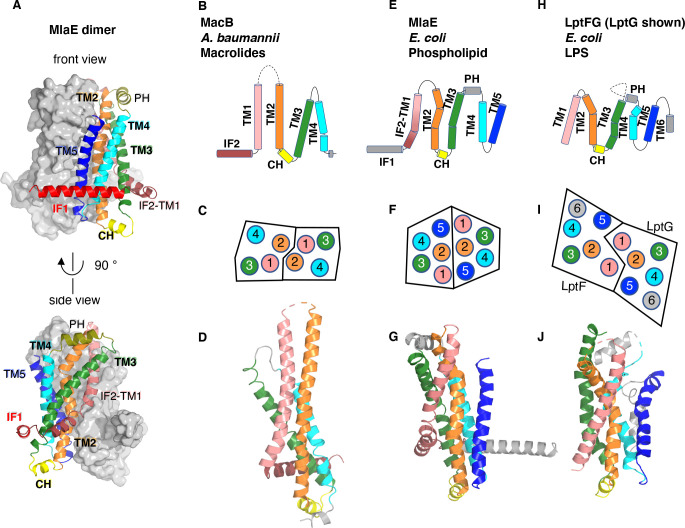
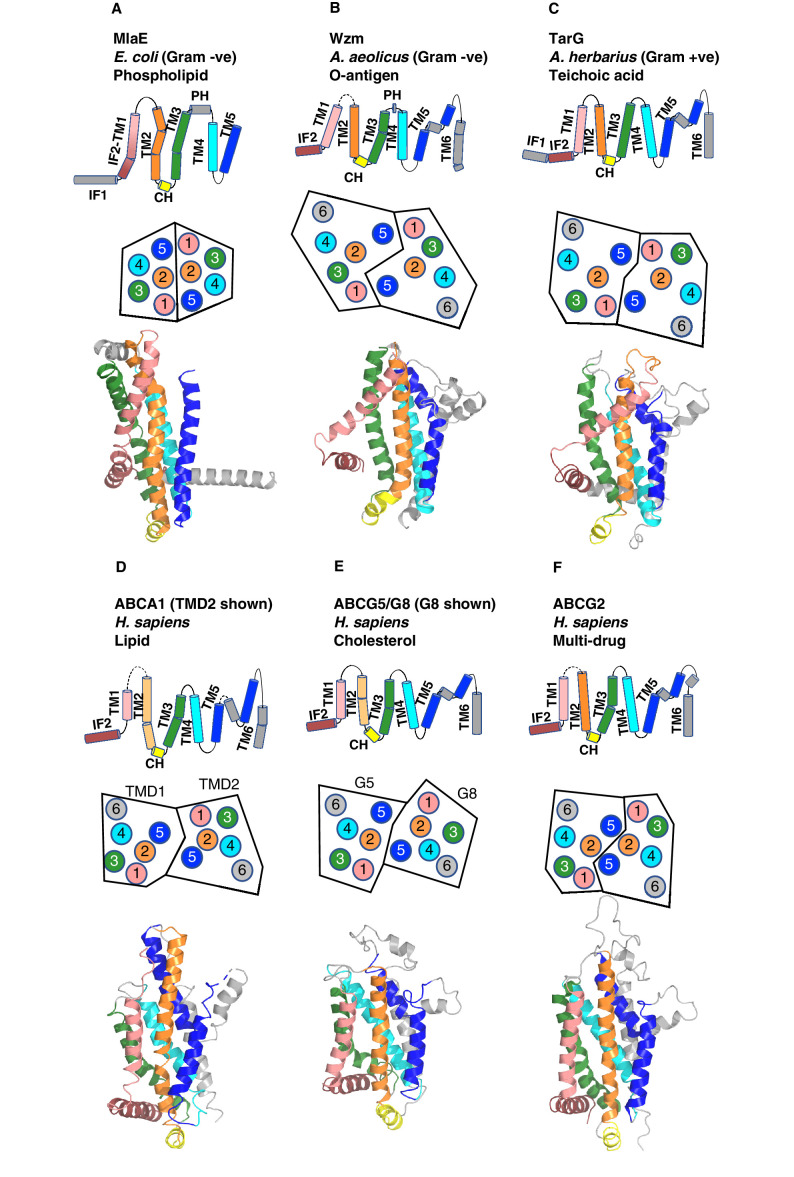
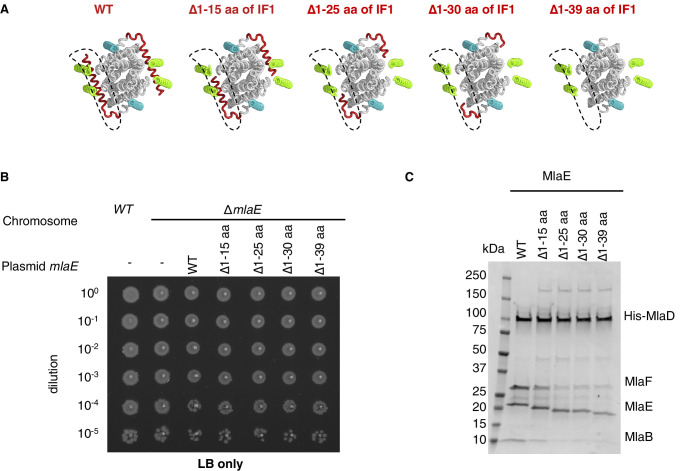
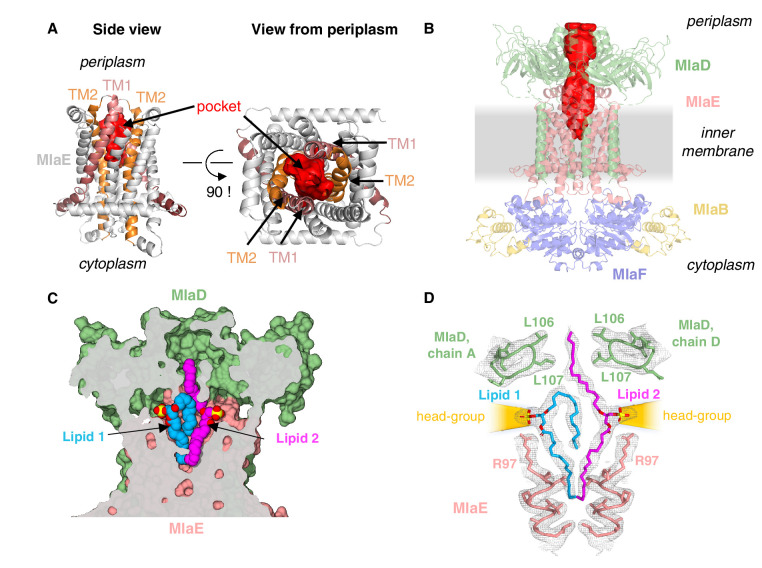
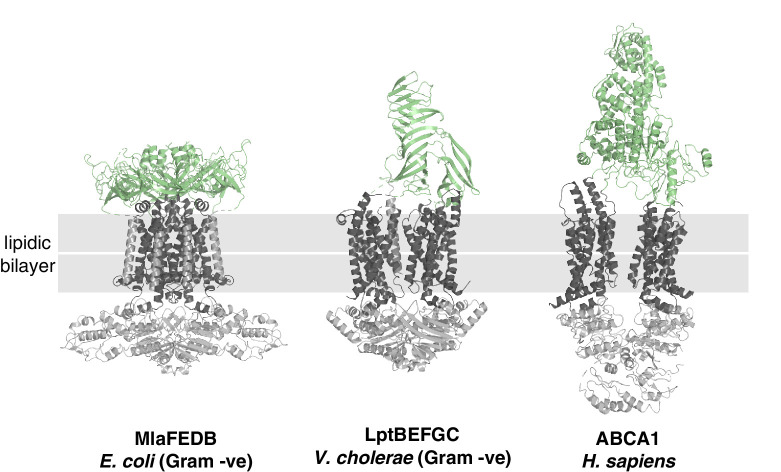
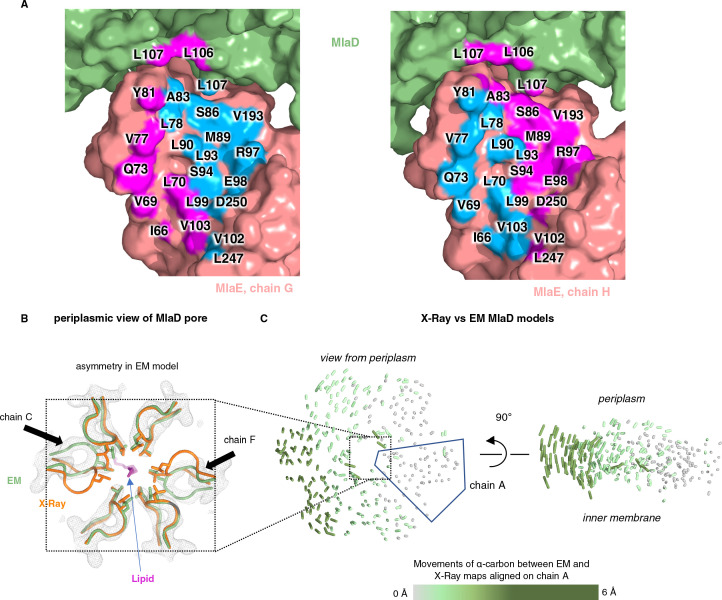
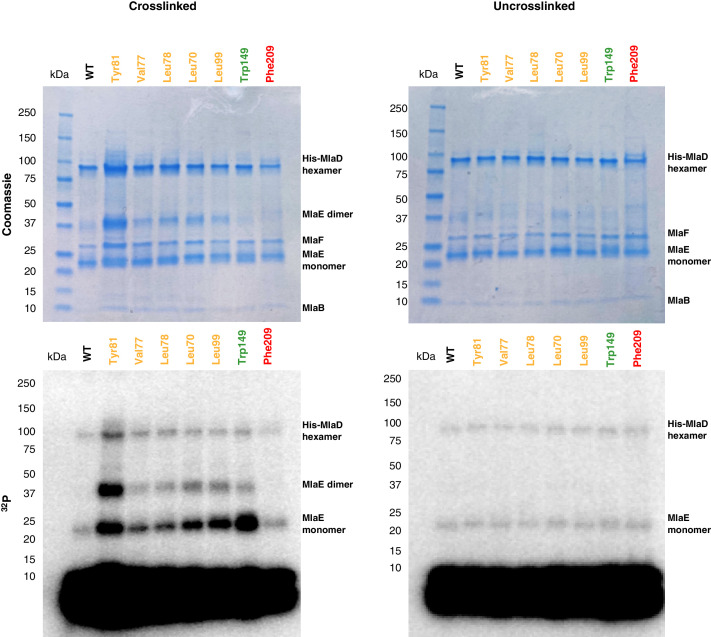
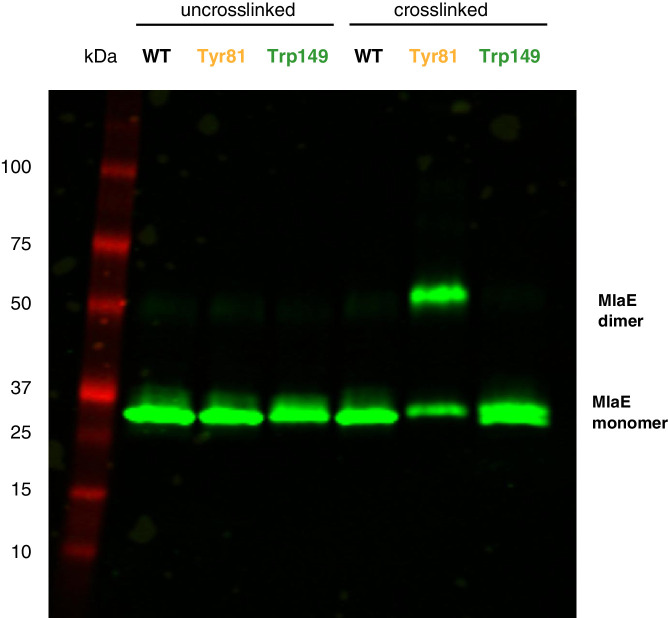
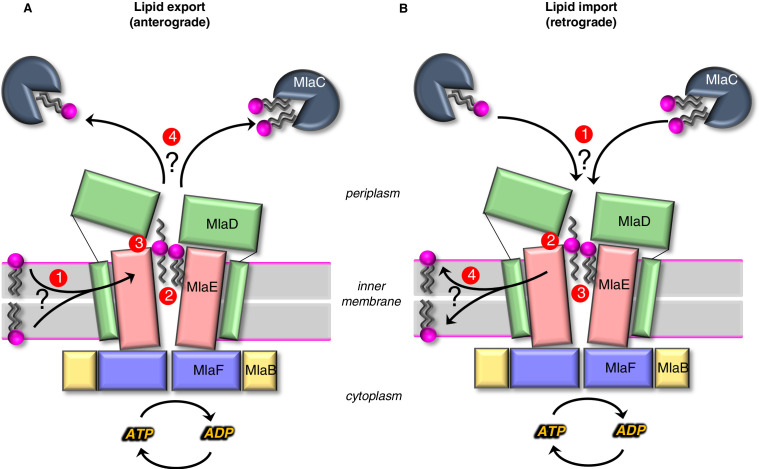
References
Publication types
MeSH terms
Substances
Grants and funding
- R00GM112982/NH/NIH HHS/United States
- P41 GM103311/GM/NIGMS NIH HHS/United States
- P41 GM103310/GM/NIGMS NIH HHS/United States
- T32 AI007180/AI/NIAID NIH HHS/United States
- R00 GM112982/GM/NIGMS NIH HHS/United States
- DFS-20-16/DRCRF/Damon Runyon Cancer Research Foundation/United States
- S10 OD019994/OD/NIH HHS/United States
- U24 GM129547/GM/NIGMS NIH HHS/United States
- T32 GM088118/GM/NIGMS NIH HHS/United States
- R35GM128777/NH/NIH HHS/United States
- T32 GM088118/NH/NIH HHS/United States
- R35 GM128777/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
