

Structural and mechanistic basis of the EMC-dependent biogenesis of distinct transmembrane clients
- PMID: 33236988
- PMCID: PMC7785296
- DOI: 10.7554/eLife.62611
Structural and mechanistic basis of the EMC-dependent biogenesis of distinct transmembrane clients
Abstract
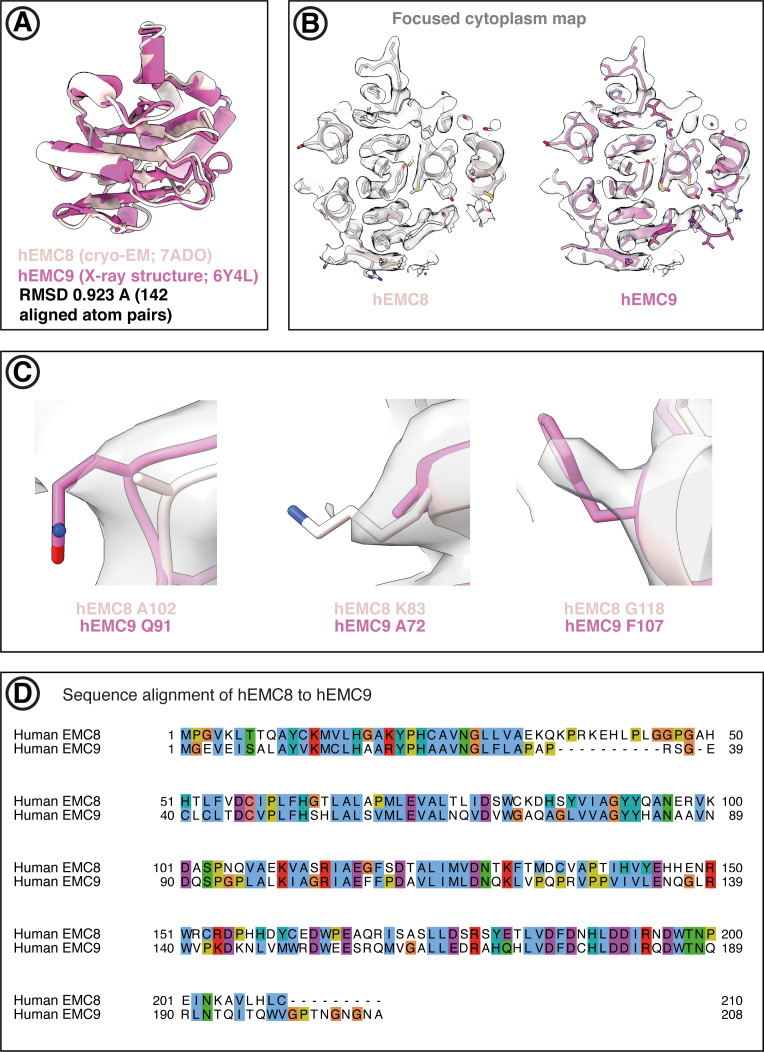
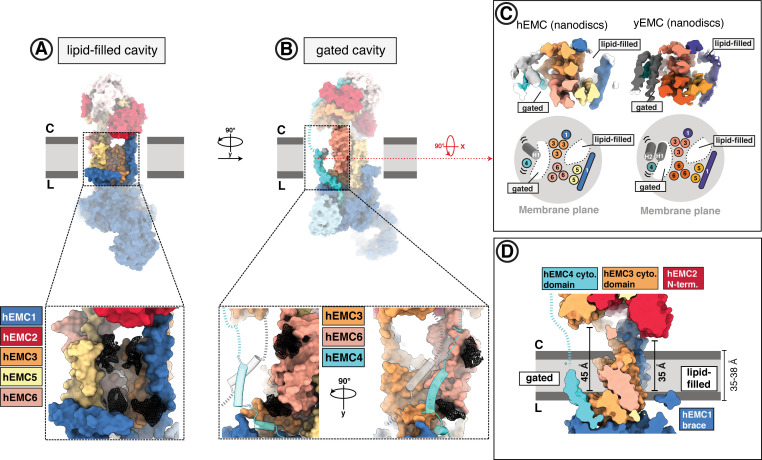
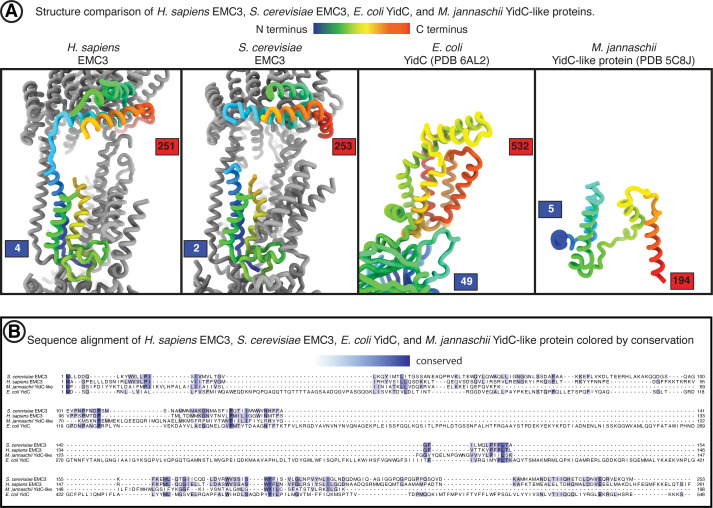
Membrane protein biogenesis in the endoplasmic reticulum (ER) is complex and failure-prone. The ER membrane protein complex (EMC), comprising eight conserved subunits, has emerged as a central player in this process. Yet, we have limited understanding of how EMC enables insertion and integrity of diverse clients, from tail-anchored to polytopic transmembrane proteins. Here, yeast and human EMC cryo-EM structures reveal conserved intricate assemblies and human-specific features associated with pathologies. Structure-based functional studies distinguish between two separable EMC activities, as an insertase regulating tail-anchored protein levels and a broader role in polytopic membrane protein biogenesis. These depend on mechanistically coupled yet spatially distinct regions including two lipid-accessible membrane cavities which confer client-specific regulation, and a non-insertase EMC function mediated by the EMC lumenal domain. Our studies illuminate the structural and mechanistic basis of EMC's multifunctionality and point to its role in differentially regulating the biogenesis of distinct client protein classes.
Keywords: EMC; S. cerevisiae; cell biology; chaperone holdase; electron microscopy; endoplasmic reticulum; human; insertase; membrane protein biogenesis; molecular biophysics; structural biology.
Plain language summary
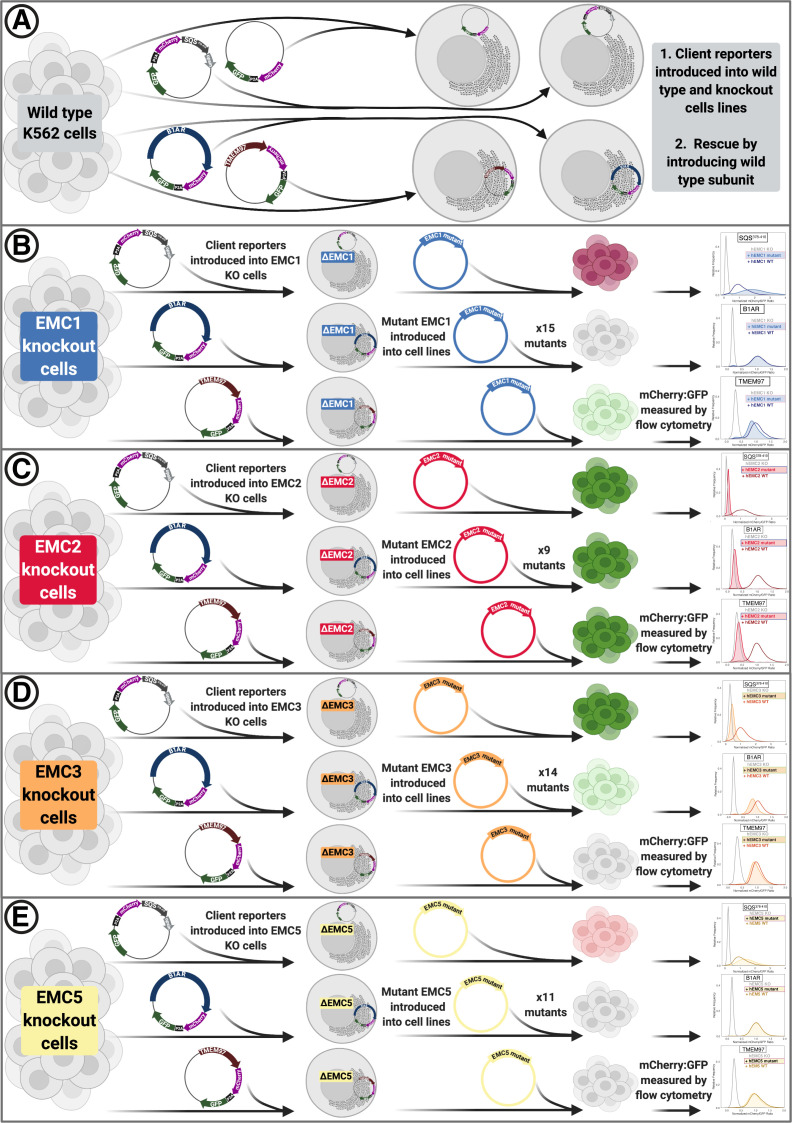
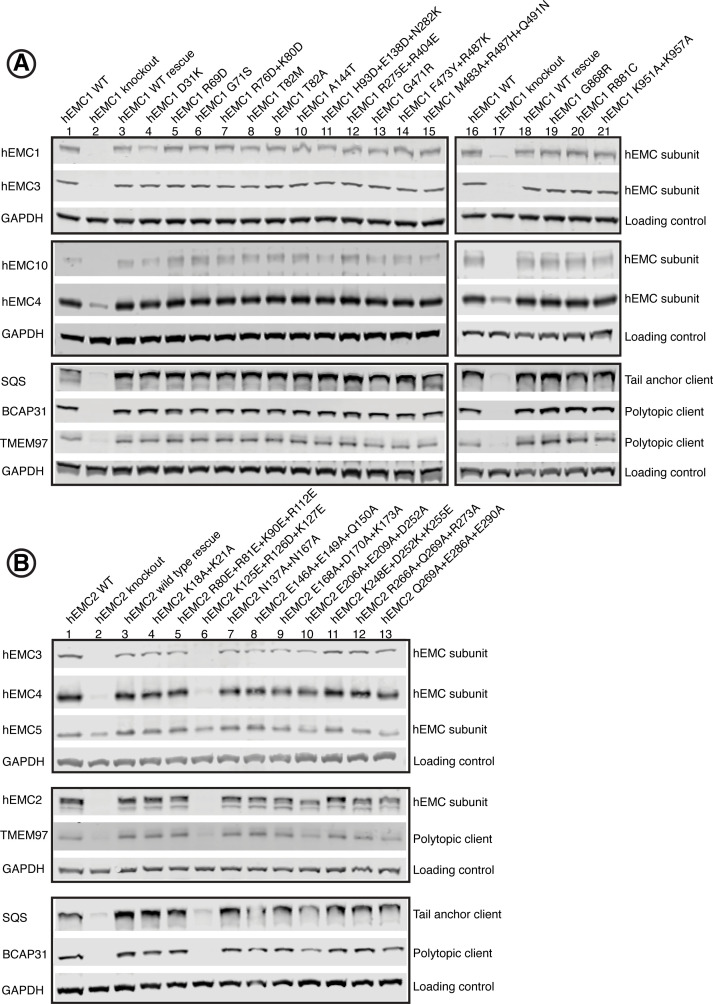
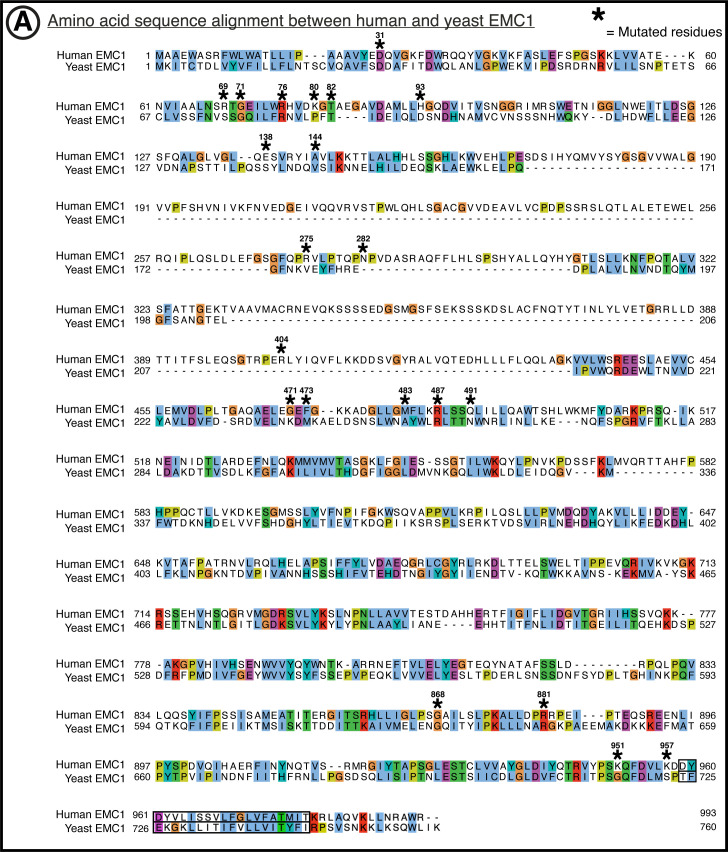
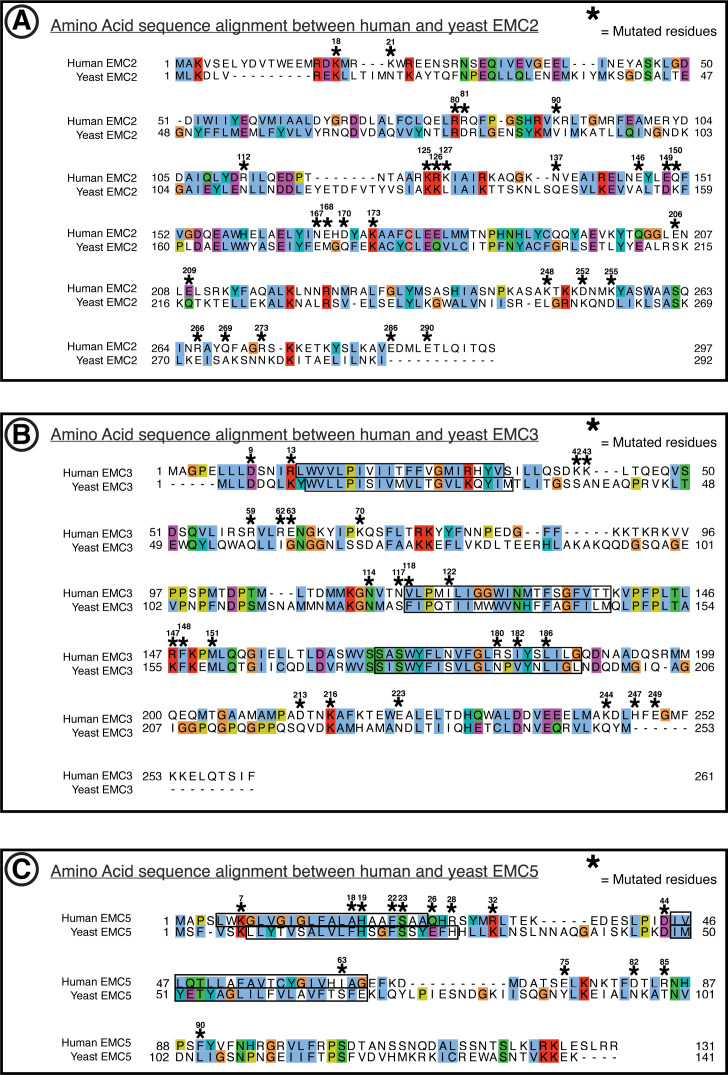
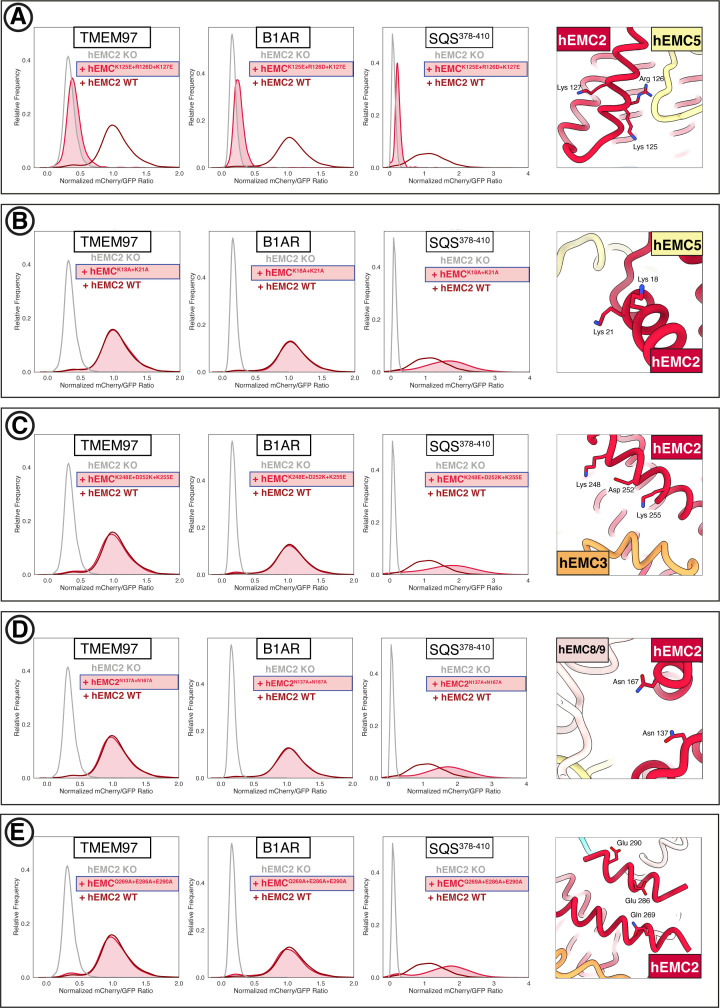
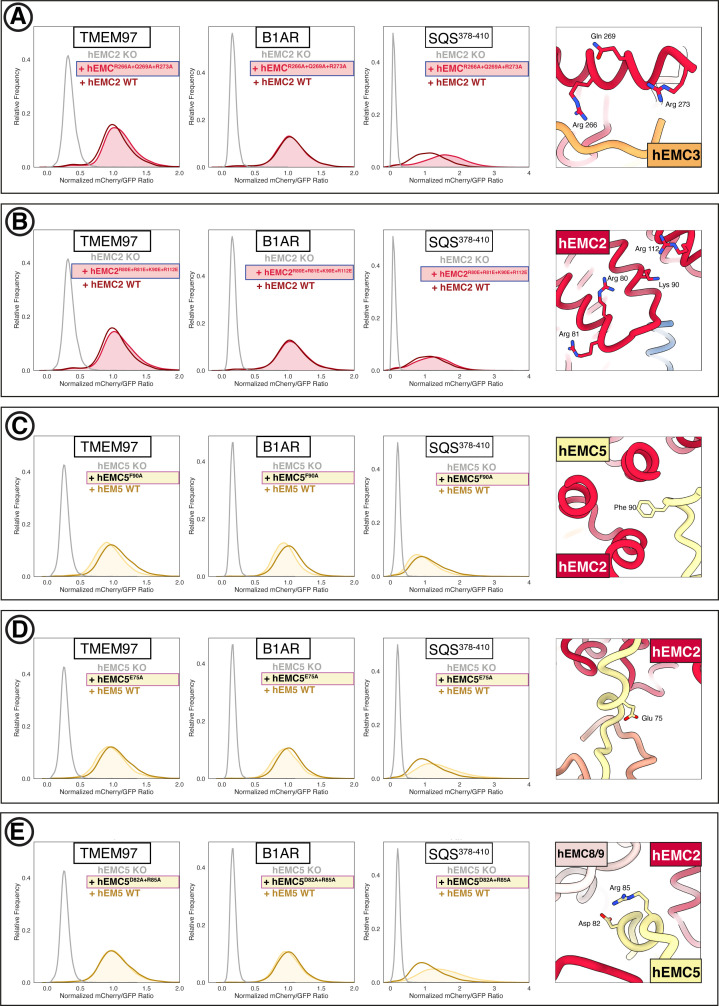
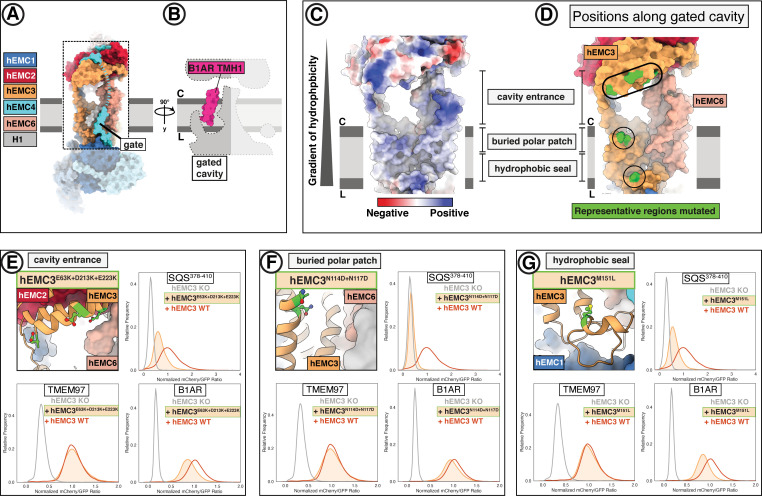
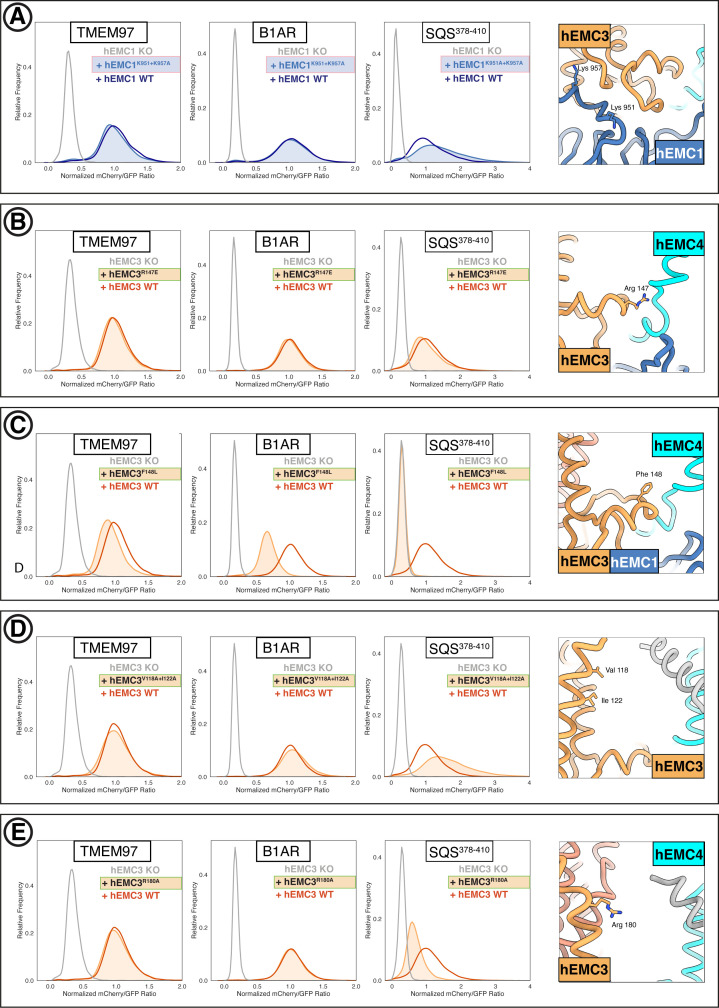
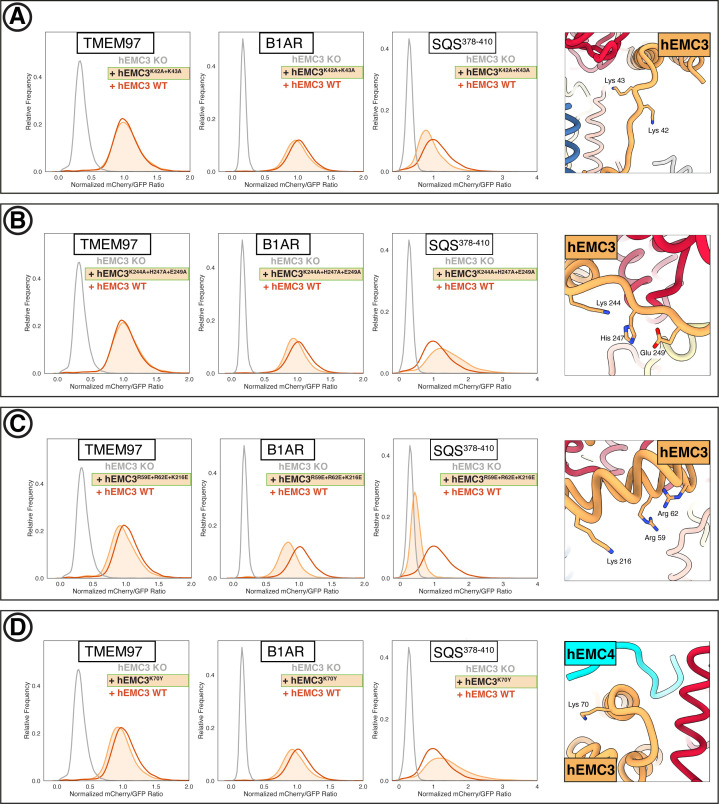
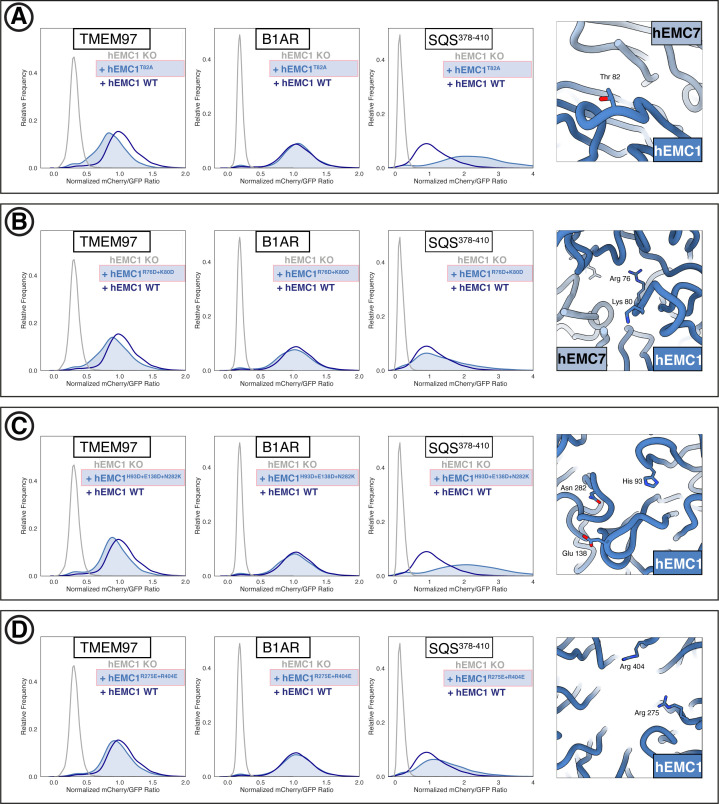
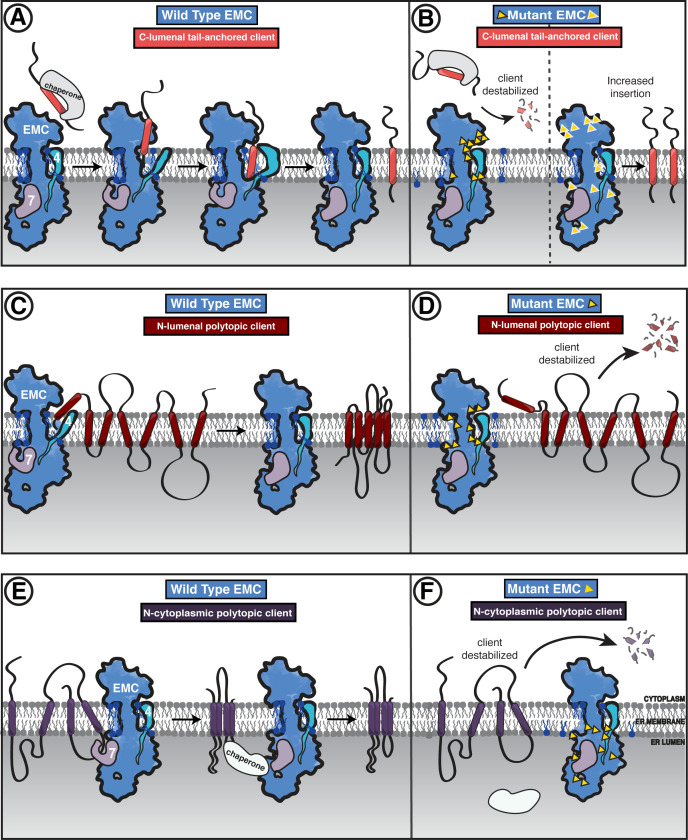
Cells are surrounded and contained by a plasma membrane consisting of a double layer of fats and proteins. These proteins monitor and facilitate the movement of food, oxygen and messages in and out of the cell, and help neighboring cells communicate. Membrane proteins are manufactured in a cell compartment called the endoplasmic reticulum. Cellular machines called ribosomes visit this compartment’s membrane to manufacture proteins that need to be secreted or embedded into the cell’s membranes. As these proteins are made, they are pulled into the endoplasmic reticulum so they can be folded correctly and inserted in the membrane. A cellular machine in this compartment’s membrane that aids this process is the endoplasmic reticulum membrane protein complex (EMC). Many steps can go wrong during protein assembly, so to control protein quality, the EMC has to accommodate the variety of complex physical features that proteins can have. To explore the activity of the EMC, Miller-Vedam, Bräuning, Popova et al. studied the normal structure of the EMC in both yeast and human cells grown in the lab. These snapshots of the complex in different species had a lot in common, including how the complex was arranged within and around the membrane. Next, Miller-Vedam, Bräuning, Popova et al. generated 50 mutant versions of the EMC in human cells to determine how changing different parts of the complex affected the production of three proteins that rely on the EMC to fold correctly. These proteins were an enzyme called squalene synthase, a signaling protein called the beta adrenergic receptor and sigma intracellular receptor 2, a protein involved in the regulation of cholesterol levels. Mutations in the section of the EMC outside of the endoplasmic reticulum, within the main cellular compartment, negatively impacted the stability of squalene synthase. This section of the EMC provides a platform where proteins can associate before entering the membrane. The part of EMC that spans the membrane contains both a fat-filled cavity and a cavity with a ‘door’ that is either open or closed. Mutations in this section disrupted the insertion of both squalene synthase and the beta adrenergic receptor into the membrane, a role performed by the cavity with the door. The specific role of the fat-filled cavity is still not fully understood, but a mutation affecting this cavity disrupts the correct production of all three proteins studied. The largest section of the complex, which sits inside the endoplasmic reticulum, protected proteins as they folded, ensuring they were not destroyed for being folded incorrectly before they were fully formed. Mutations in this part of the EMC negatively impacted the stability of sigma intracellular receptor 2 without negatively affecting the other proteins. This molecular dissection of the activity of the EMC provides insights into how membrane proteins are manufactured, stabilized, coordinated, and monitored for quality. These findings could contribute towards the development of new treatments for certain congenital diseases. For example, cystic fibrosis, retinitis pigmentosa, and Charcot-Marie-Tooth disease are all thought to be caused by mutations within membrane proteins that require the EMC during their production.
© 2020, Miller-Vedam et al.
Conflict of interest statement
LM, BB, KP, NS, JB, JP, EB, NS, MS, RS, CC, BS, AF, JW No competing interests declared
Figures
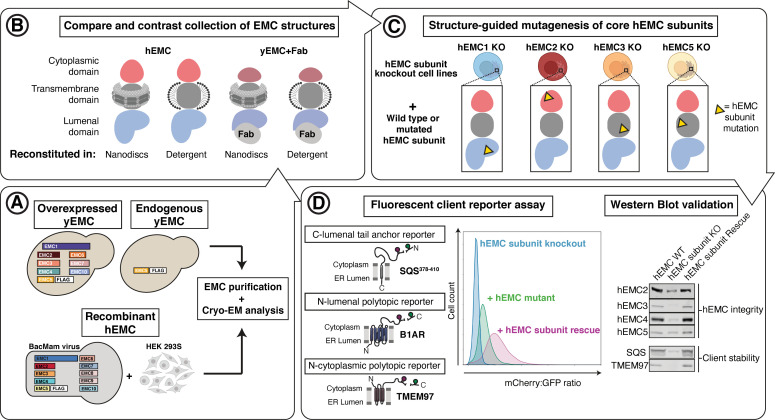
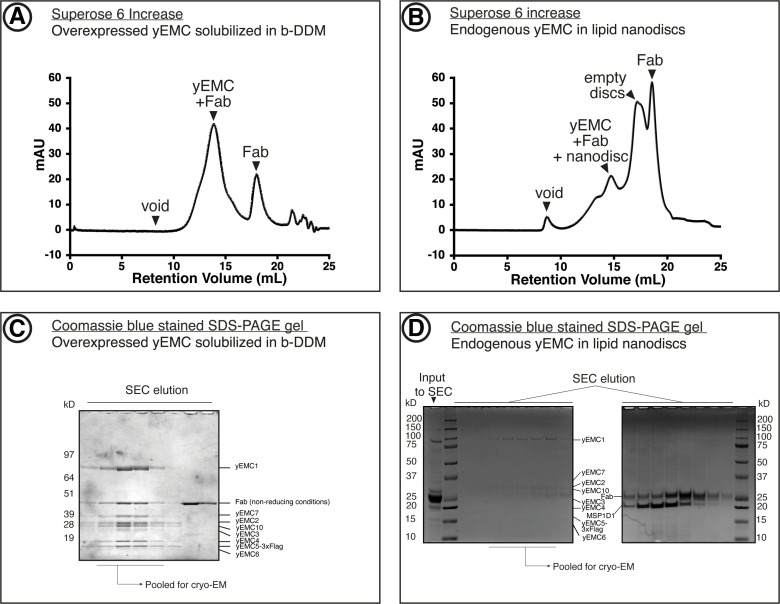
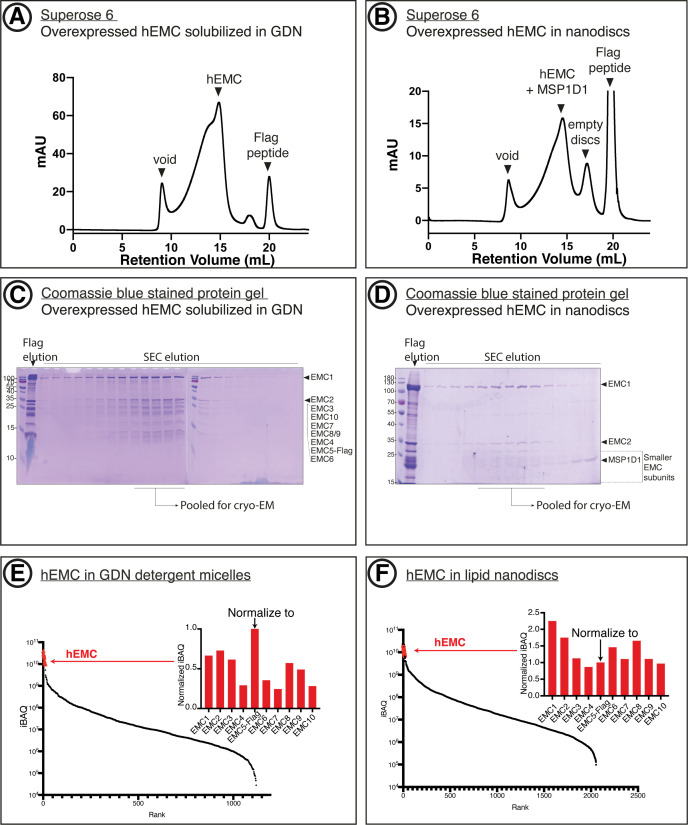
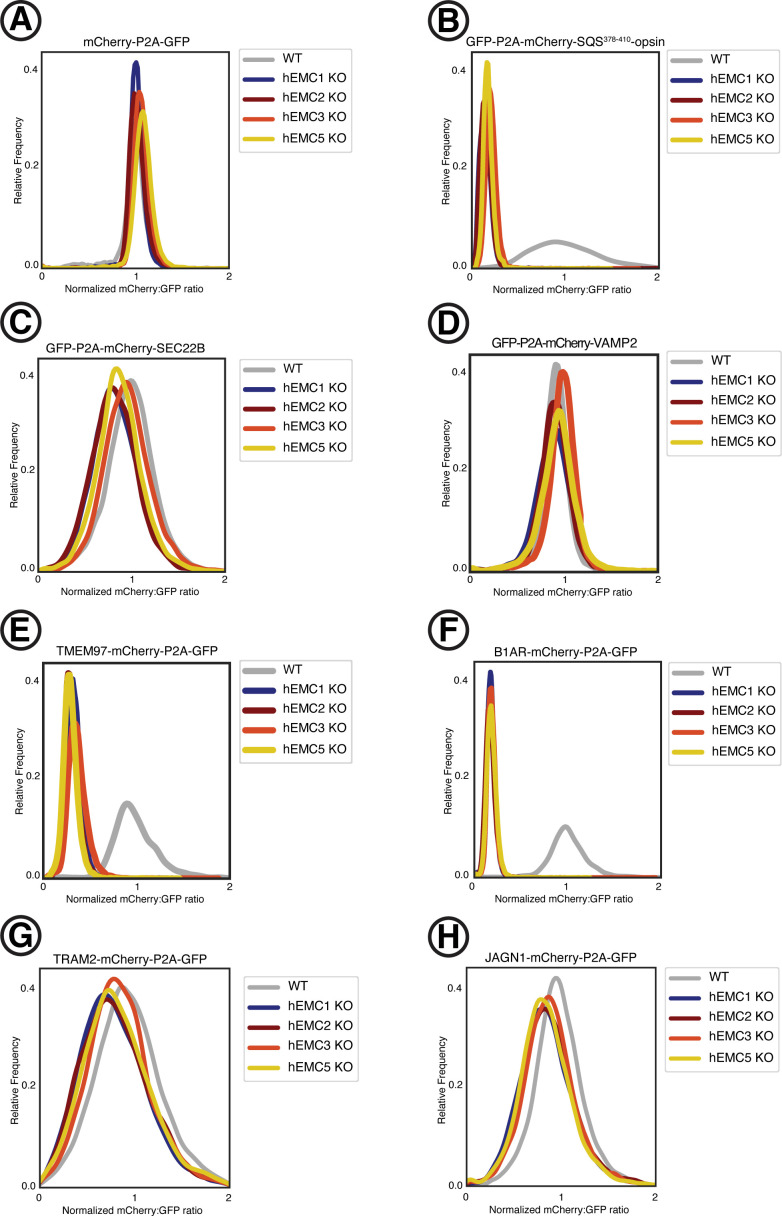

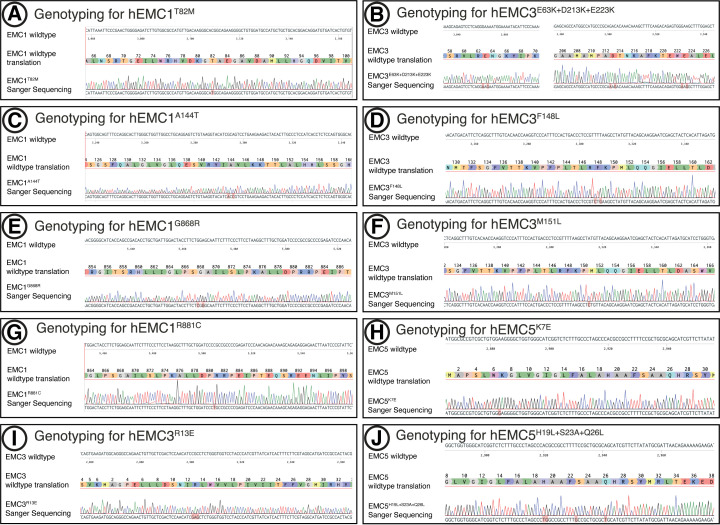
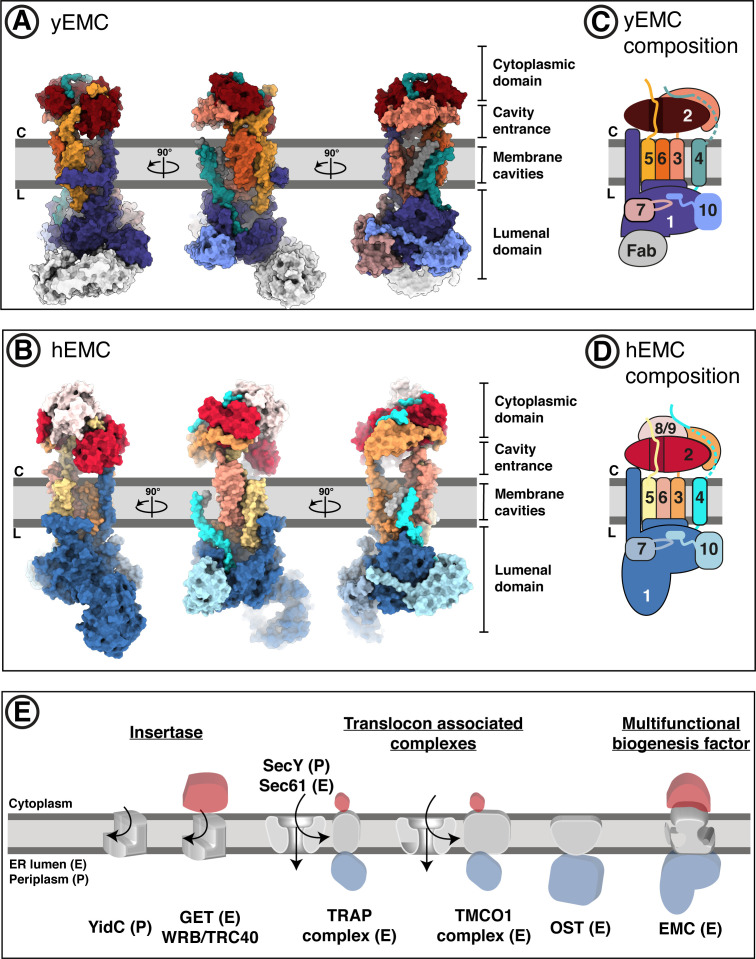
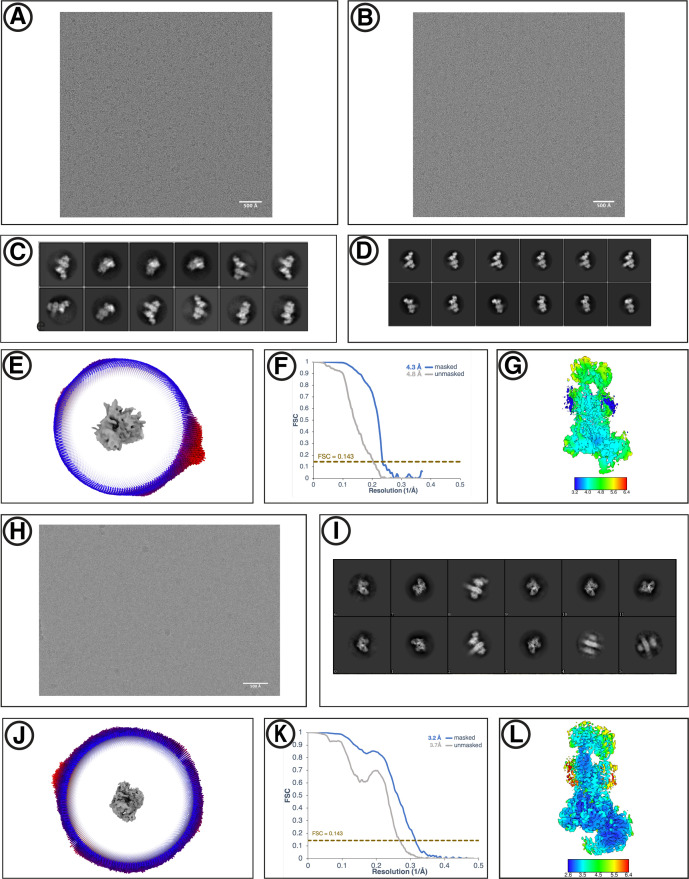
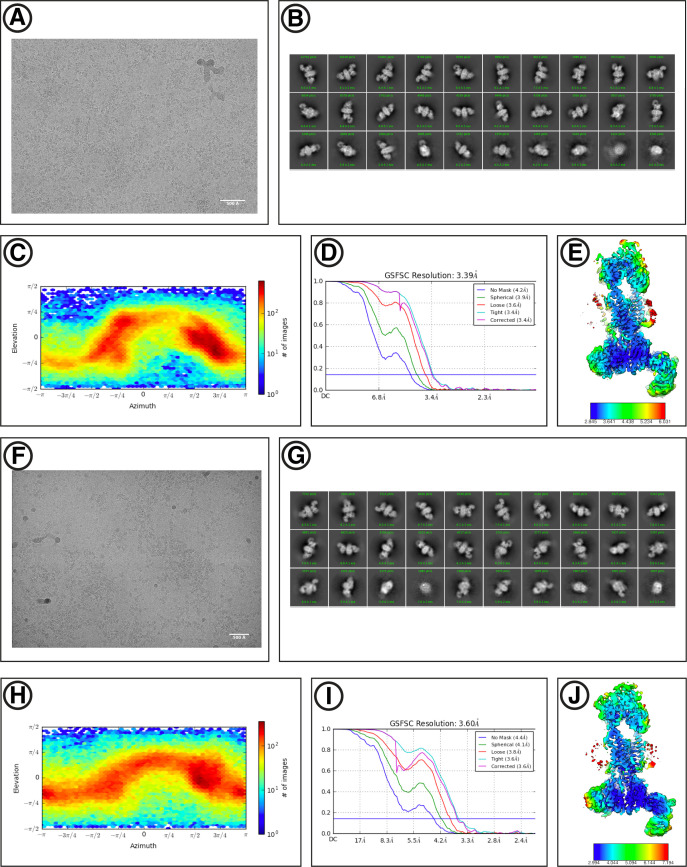
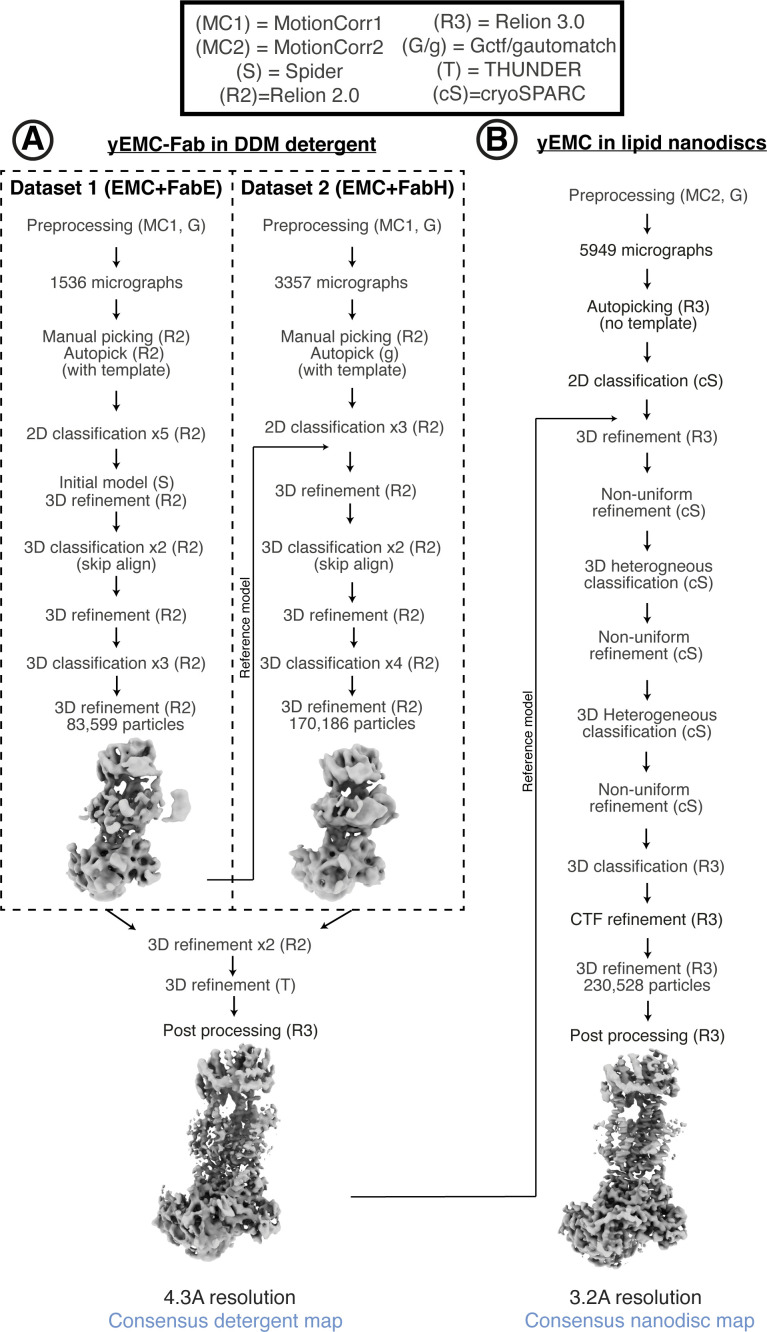
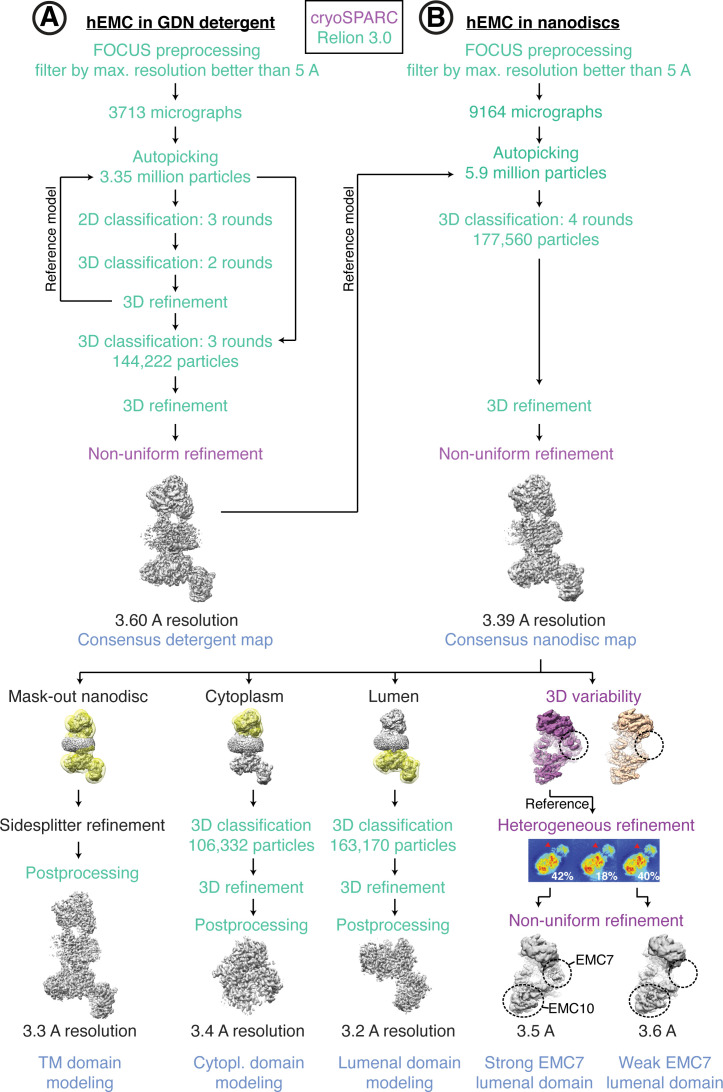
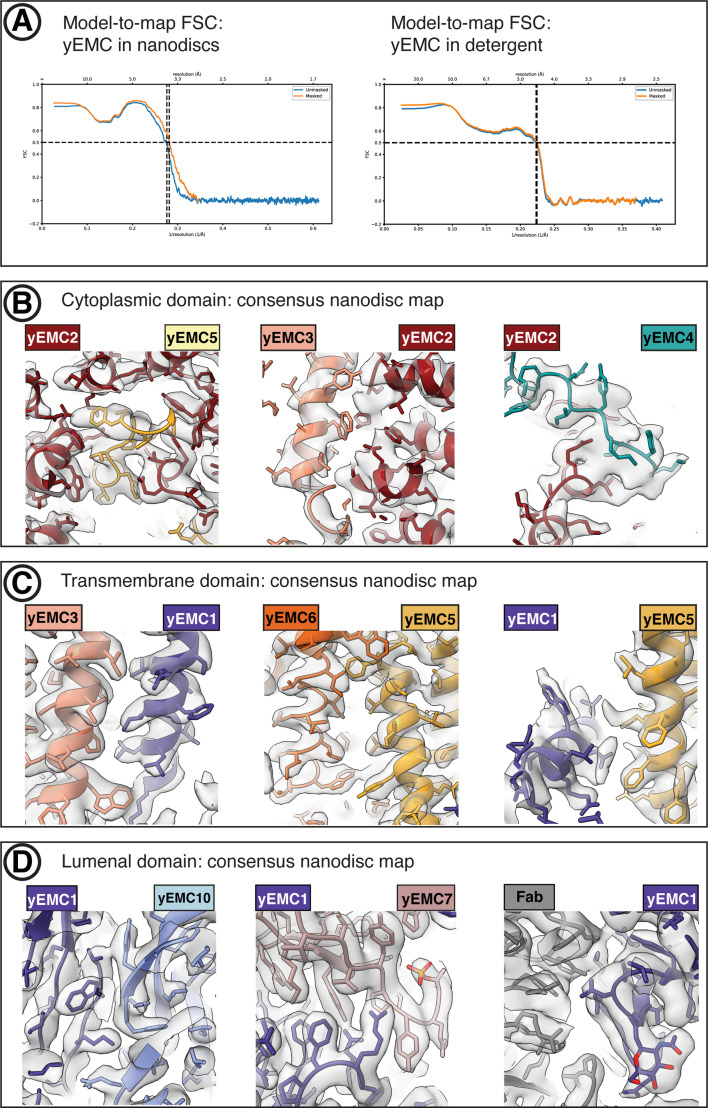
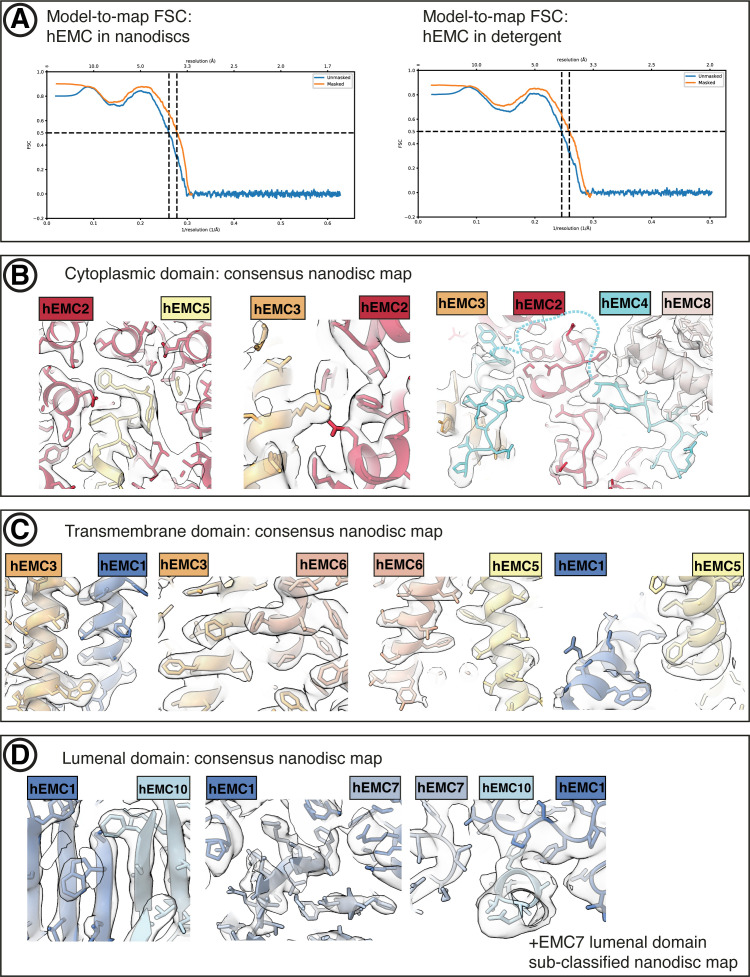
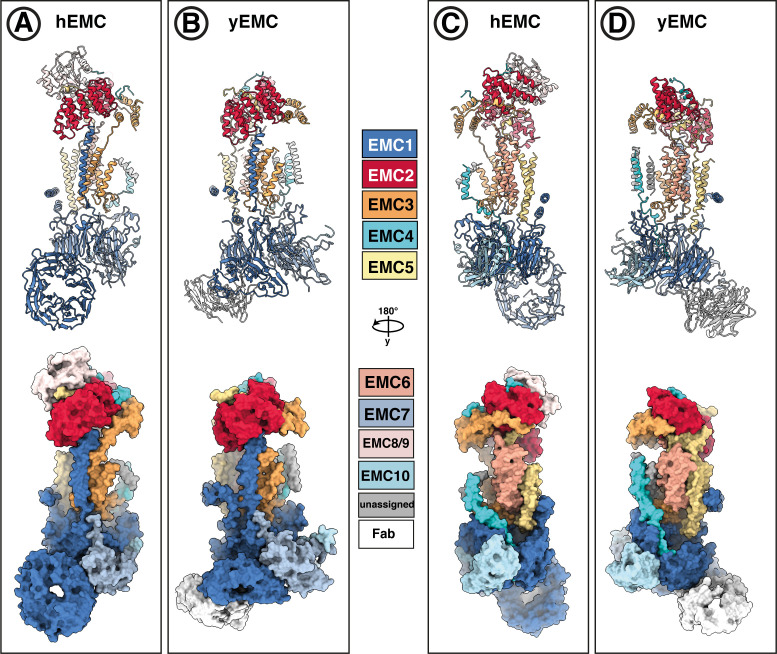
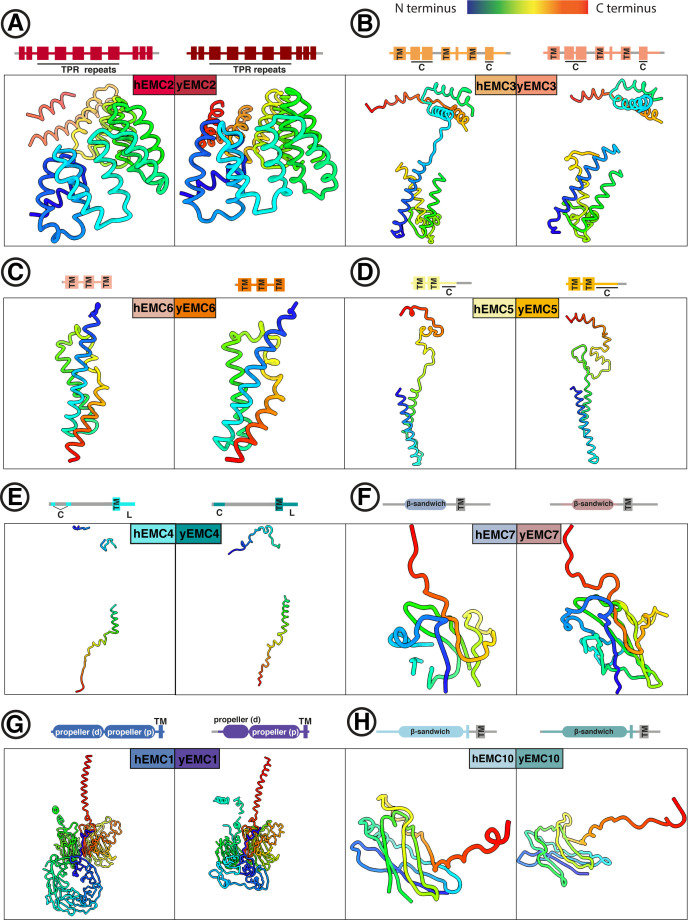
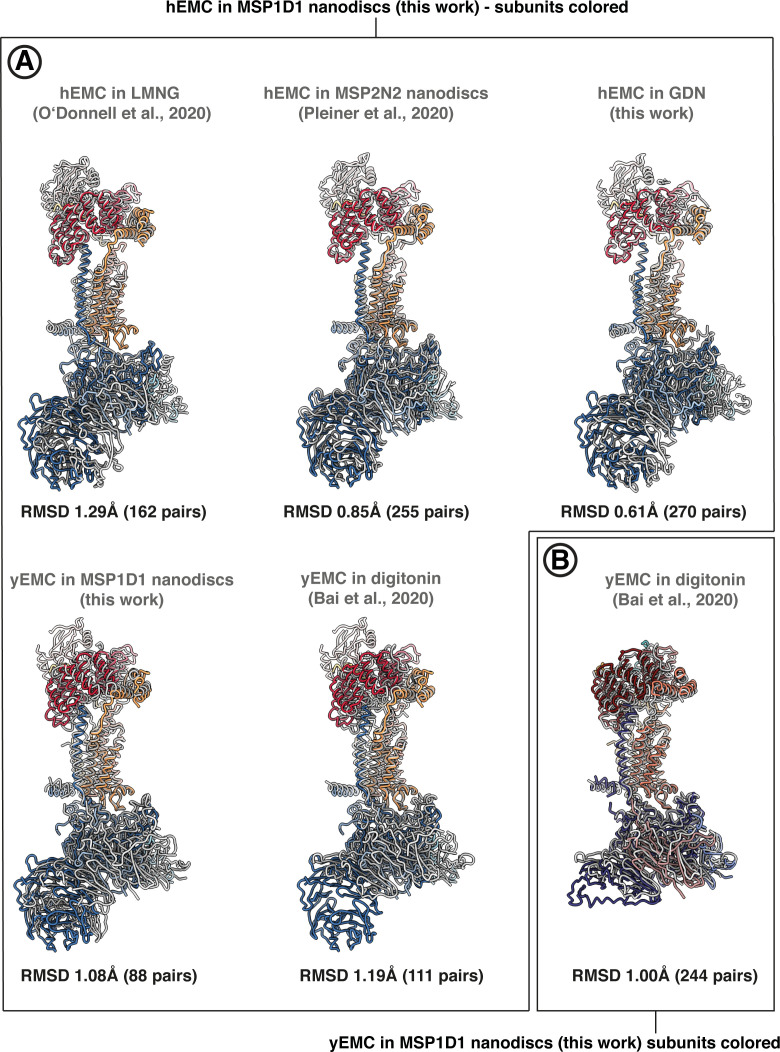
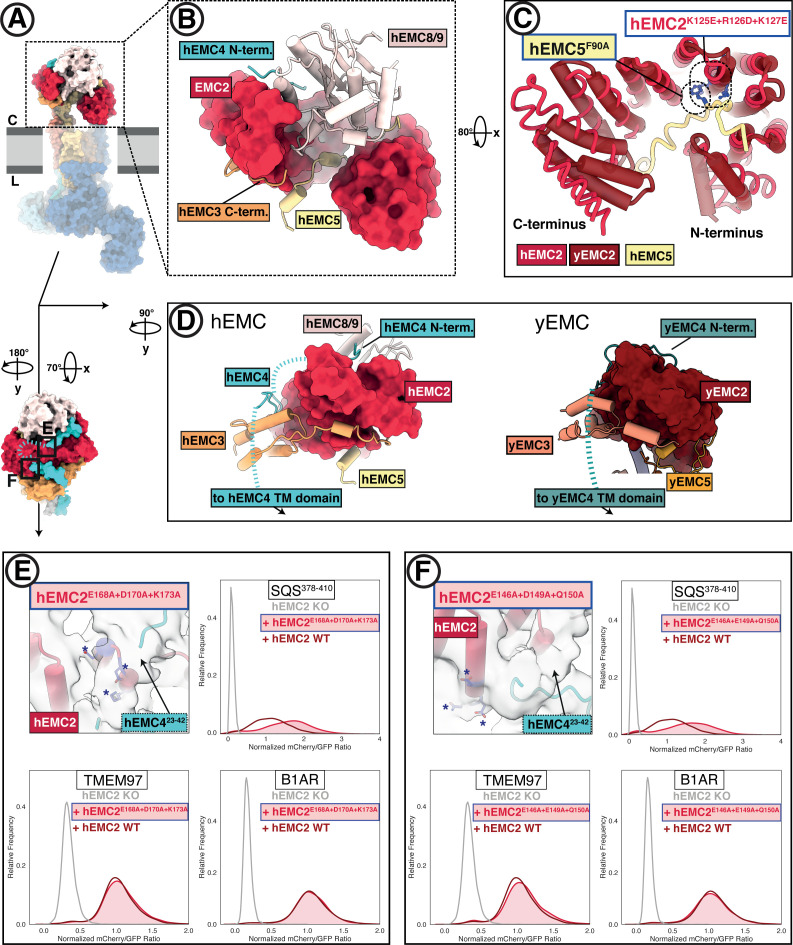
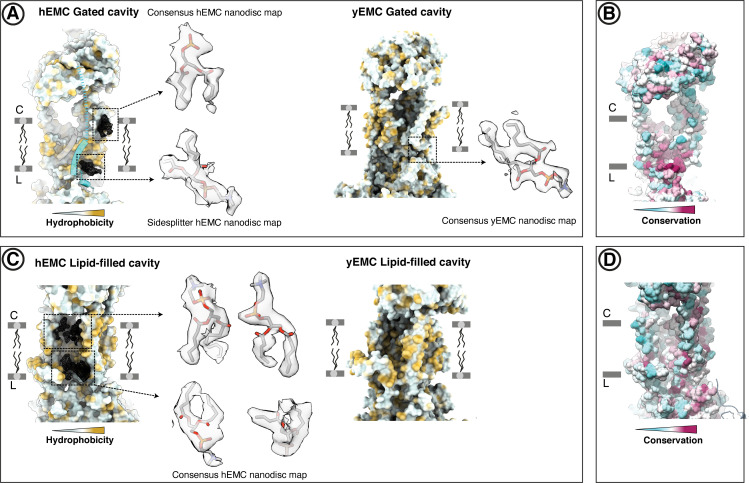
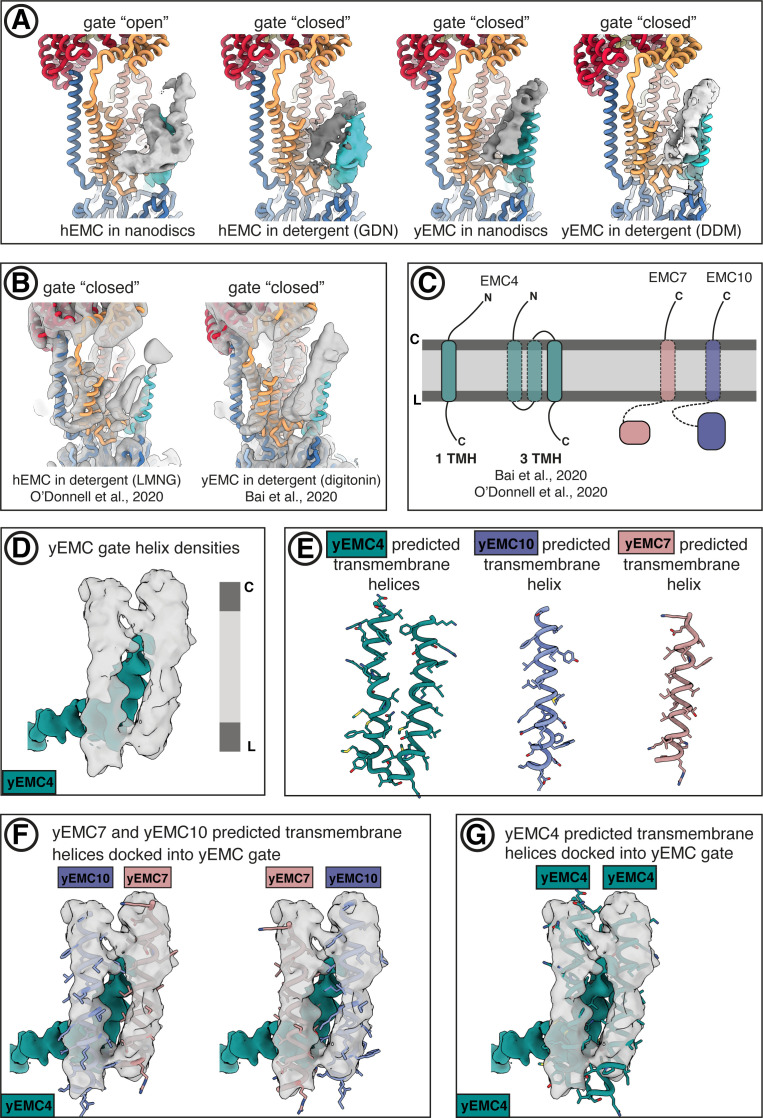
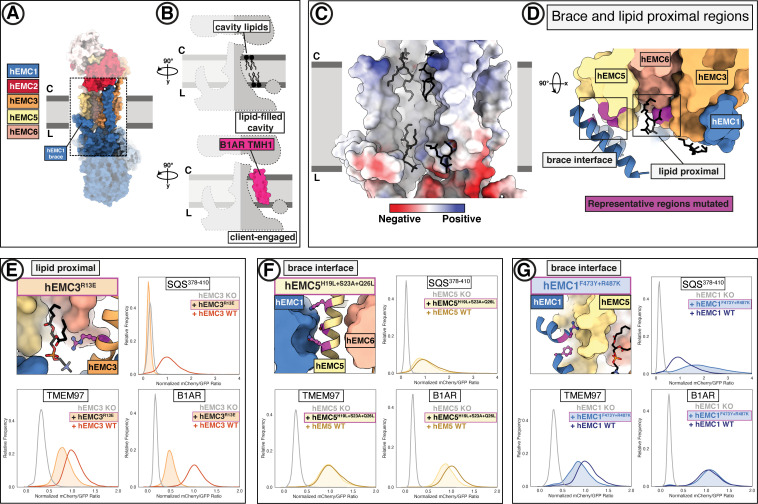
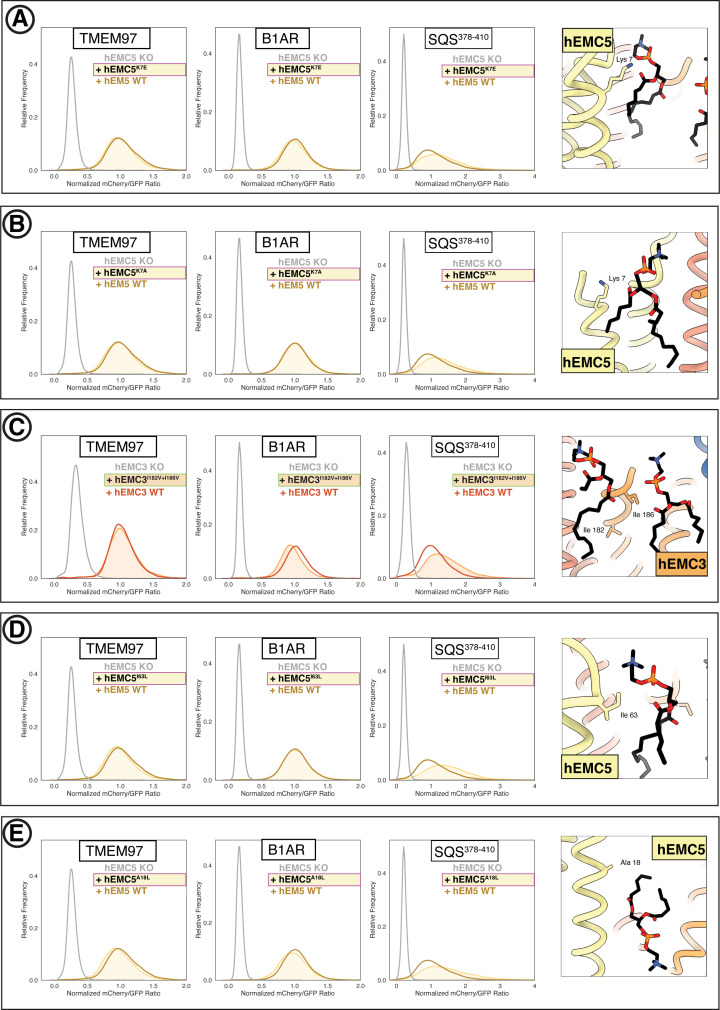
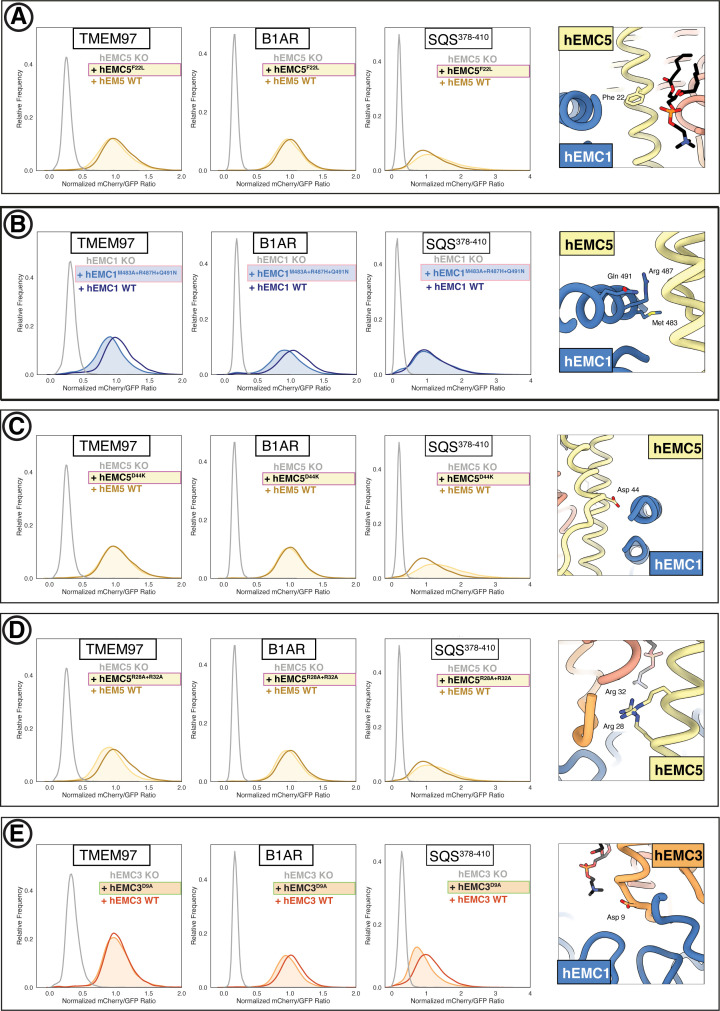
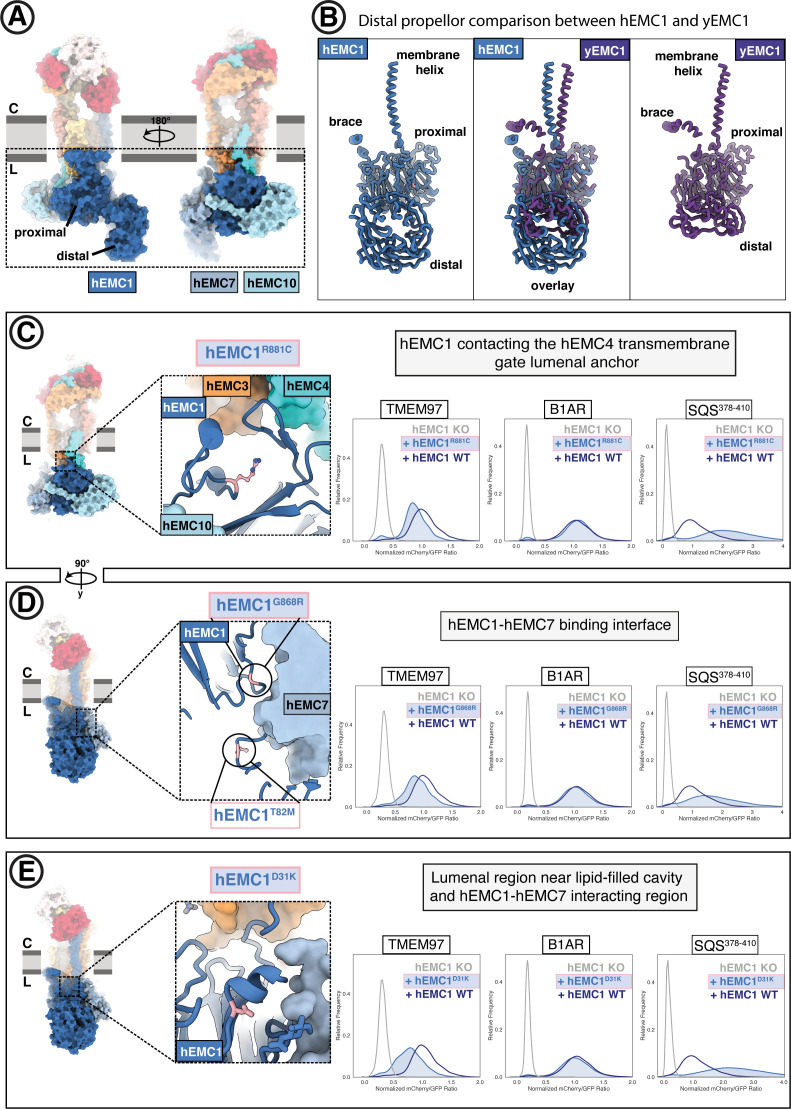
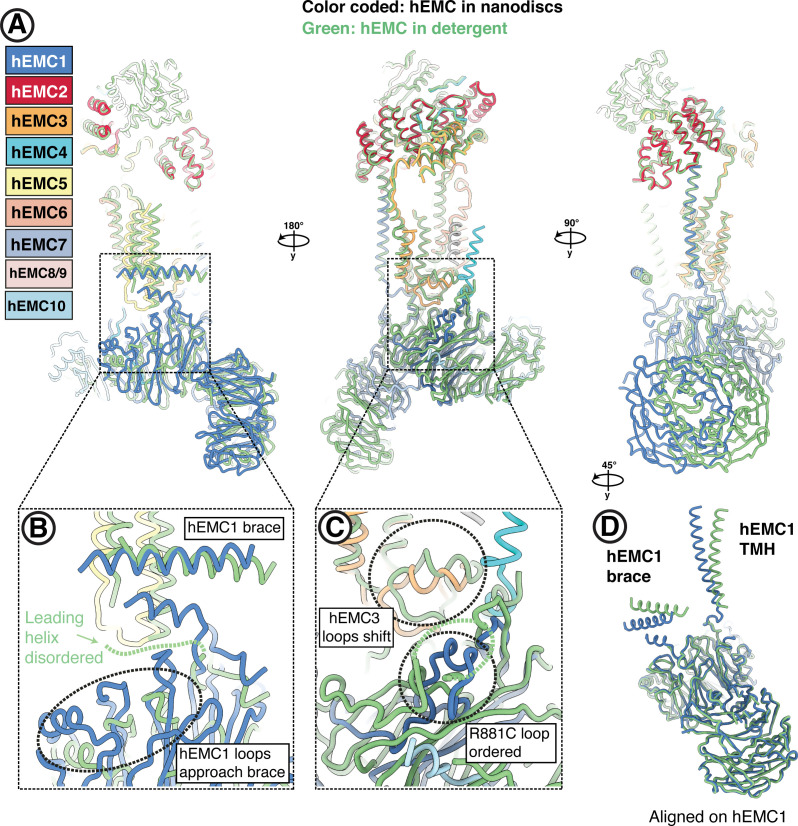
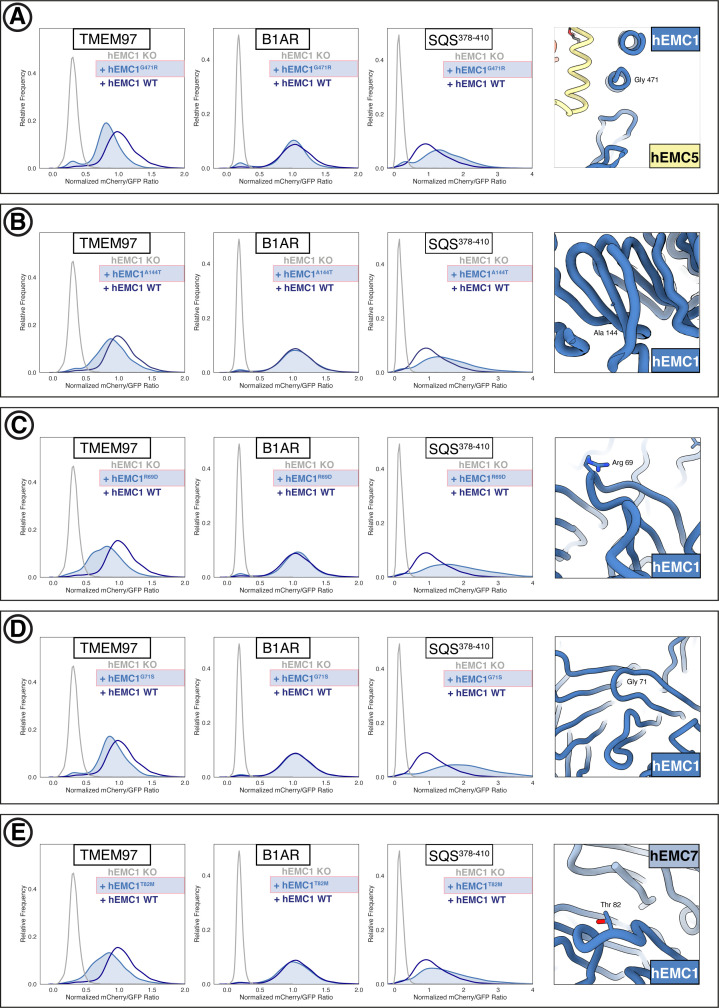

Similar articles
-
An ER translocon for multi-pass membrane protein biogenesis.Elife. 2020 Aug 21;9:e56889. doi: 10.7554/eLife.56889. Elife. 2020. PMID: 32820719 Free PMC article.
-
The Get1/2 transmembrane complex is an endoplasmic-reticulum membrane protein insertase.Nature. 2014 Aug 28;512(7515):441-4. doi: 10.1038/nature13471. Epub 2014 Jul 20. Nature. 2014. PMID: 25043001 Free PMC article.
-
Structural insights into human EMC and its interaction with VDAC.Aging (Albany NY). 2024 Mar 15;16(6):5501-5525. doi: 10.18632/aging.205660. Epub 2024 Mar 15. Aging (Albany NY). 2024. PMID: 38517390 Free PMC article.
-
Cryo-EM structures of the endoplasmic reticulum membrane complex.FEBS J. 2022 Jan;289(1):102-112. doi: 10.1111/febs.15786. Epub 2021 Mar 6. FEBS J. 2022. PMID: 33629497 Free PMC article. Review.
-
Cryo-EM insights into tail-anchored membrane protein biogenesis in eukaryotes.Curr Opin Struct Biol. 2022 Aug;75:102428. doi: 10.1016/j.sbi.2022.102428. Epub 2022 Jul 16. Curr Opin Struct Biol. 2022. PMID: 35850079 Review.
Cited by
-
Structural biology of voltage-gated calcium channels.Channels (Austin). 2024 Dec;18(1):2290807. doi: 10.1080/19336950.2023.2290807. Epub 2023 Dec 7. Channels (Austin). 2024. PMID: 38062897 Free PMC article. Review.
-
A toolbox for systematic discovery of stable and transient protein interactors in baker's yeast.Mol Syst Biol. 2023 Feb 10;19(2):e11084. doi: 10.15252/msb.202211084. Epub 2023 Jan 18. Mol Syst Biol. 2023. PMID: 36651308 Free PMC article.
-
A Structural and Bioinformatics Investigation of a Fungal Squalene Synthase and Comparisons with Other Membrane Proteins.ACS Omega. 2022 Jun 17;7(26):22601-22612. doi: 10.1021/acsomega.2c01924. eCollection 2022 Jul 5. ACS Omega. 2022. PMID: 35811857 Free PMC article.
-
The mechanisms of integral membrane protein biogenesis.Nat Rev Mol Cell Biol. 2022 Feb;23(2):107-124. doi: 10.1038/s41580-021-00413-2. Epub 2021 Sep 23. Nat Rev Mol Cell Biol. 2022. PMID: 34556847 Free PMC article. Review.
-
Co-translational biogenesis of lipid droplet integral membrane proteins.J Cell Sci. 2022 Mar 1;135(5):jcs259220. doi: 10.1242/jcs.259220. Epub 2021 Nov 2. J Cell Sci. 2022. PMID: 34558621 Free PMC article.
References
-
- Abu-Safieh L, Alrashed M, Anazi S, Alkuraya H, Khan AO, Al-Owain M, Al-Zahrani J, Al-Abdi L, Hashem M, Al-Tarimi S, Sebai MA, Shamia A, Ray-Zack MD, Nassan M, Al-Hassnan ZN, Rahbeeni Z, Waheeb S, Alkharashi A, Abboud E, Al-Hazzaa SA, Alkuraya FS. Autozygome-guided exome sequencing in retinal dystrophy patients reveals pathogenetic mutations and novel candidate disease genes. Genome Research. 2013;23:236–247. doi: 10.1101/gr.144105.112. - DOI - PMC - PubMed
-
- Adams PD, Afonine PV, Bunkóczi G, Chen VB, Echols N, Headd JJ, Hung LW, Jain S, Kapral GJ, Grosse Kunstleve RW, McCoy AJ, Moriarty NW, Oeffner RD, Read RJ, Richardson DC, Richardson JS, Terwilliger TC, Zwart PH. The Phenix software for automated determination of macromolecular structures. Methods. 2011;55:94–106. doi: 10.1016/j.ymeth.2011.07.005. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM024485/GM/NIGMS NIH HHS/United States
- S10 OD020054/OD/NIH HHS/United States
- P30 CA082103/CA/NCI NIH HHS/United States
- R01 GM129325/GM/NIGMS NIH HHS/United States
- P41 CA196276/CA/NCI NIH HHS/United States
- 55108523/HHMI/Howard Hughes Medical Institute/United States
- GM24485/NH/NIH HHS/United States
- P50 AI150476/AI/NIAID NIH HHS/United States
- S10 OD021596/OD/NIH HHS/United States
- R37 GM024485/GM/NIGMS NIH HHS/United States
- S10 OD021741/OD/NIH HHS/United States
- 1DP2OD017690-01/NH/NIH HHS/United States
- P41 GM103311/GM/NIGMS NIH HHS/United States
- 1P41CA196276-01/NH/NIH HHS/United States
- T32 GM008284/GM/NIGMS NIH HHS/United States
- P50AI150476/NH/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
