

Compromised counterselection by FAS creates an aggressive subtype of germinal center lymphoma
- PMID: 33237303
- PMCID: PMC7694576
- DOI: 10.1084/jem.20201173
Compromised counterselection by FAS creates an aggressive subtype of germinal center lymphoma
Abstract
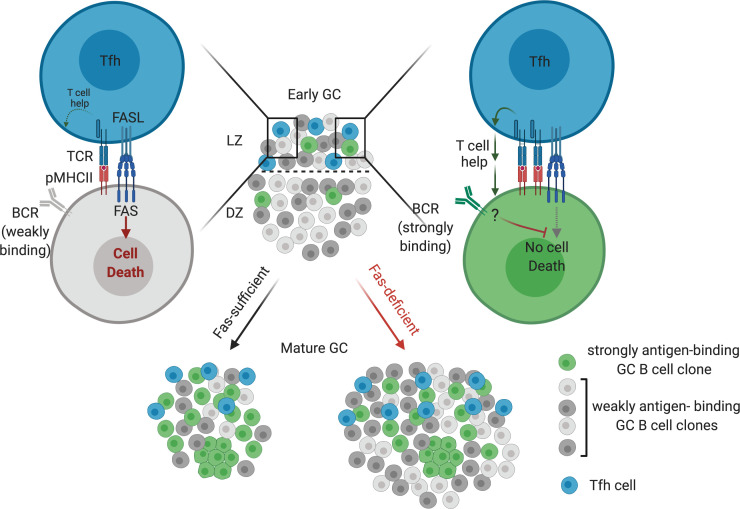
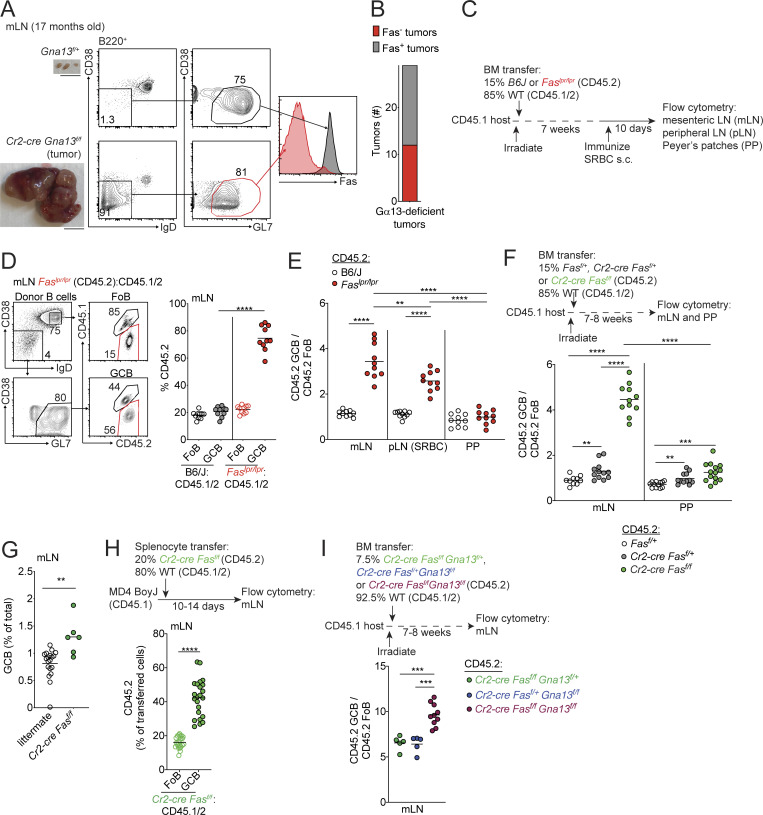
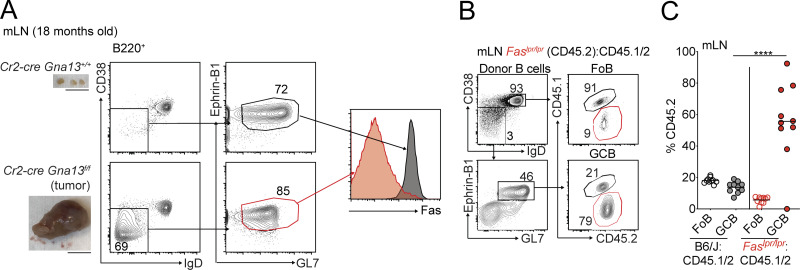
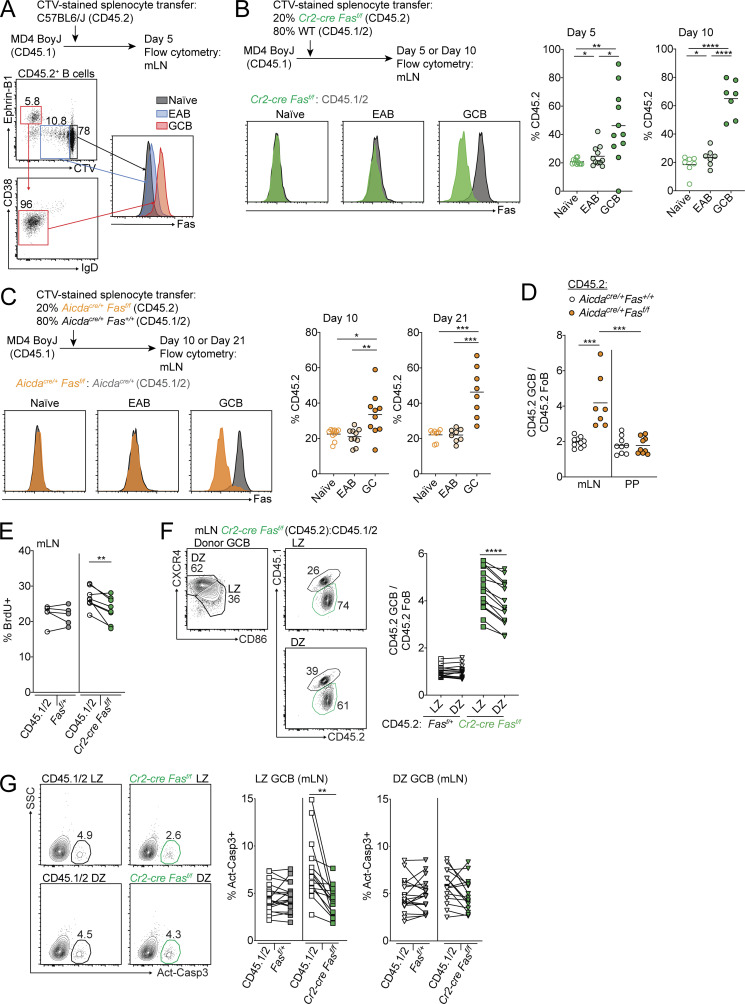
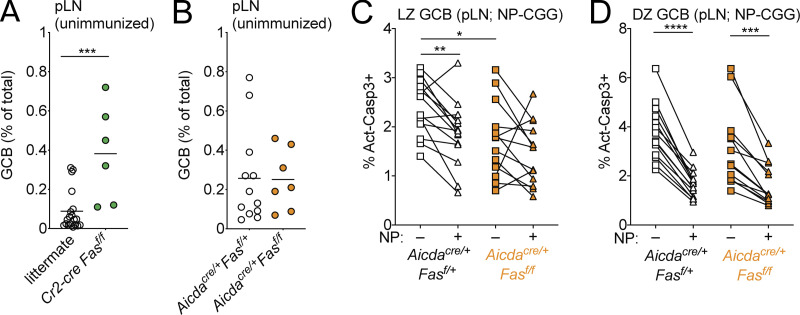
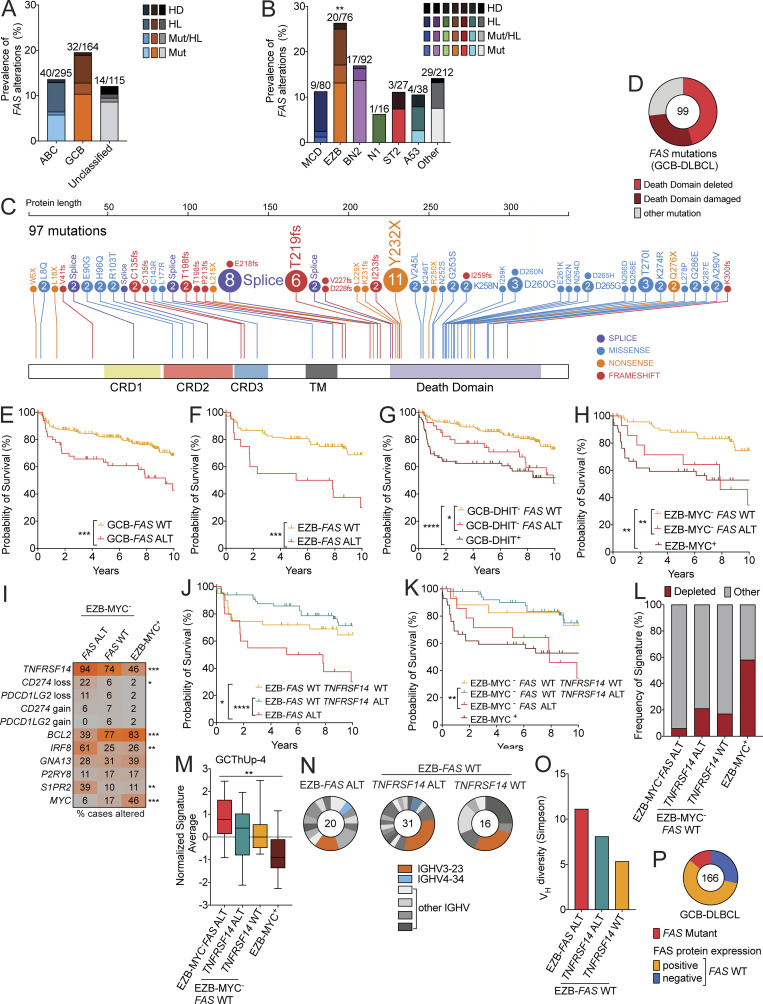
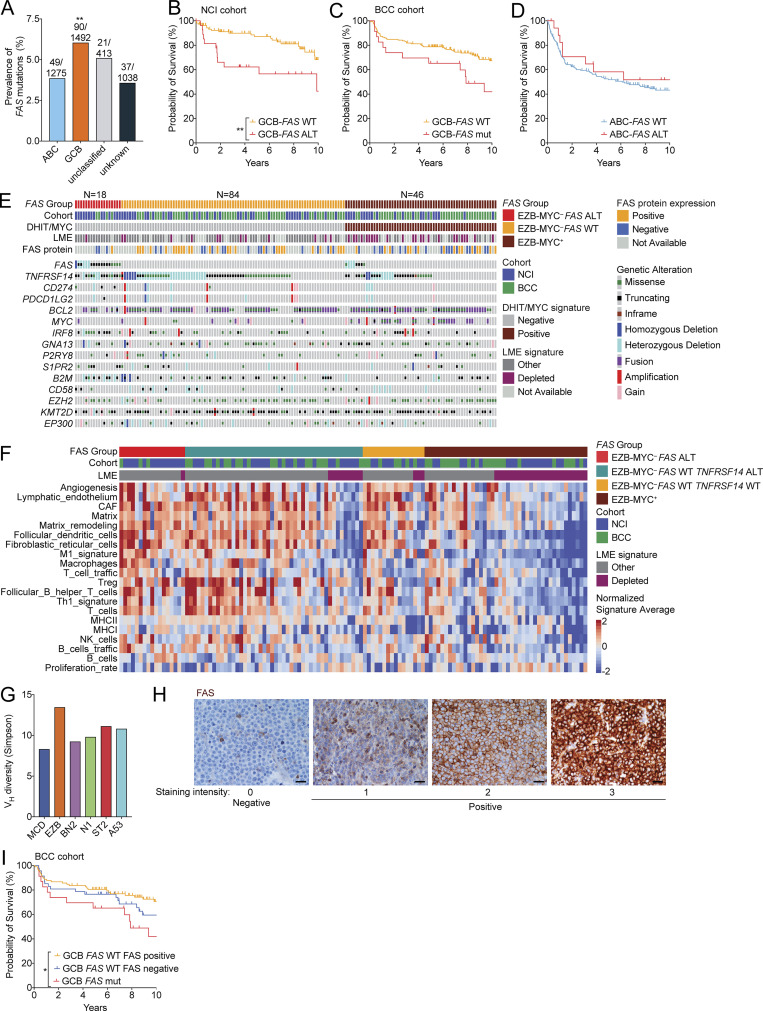
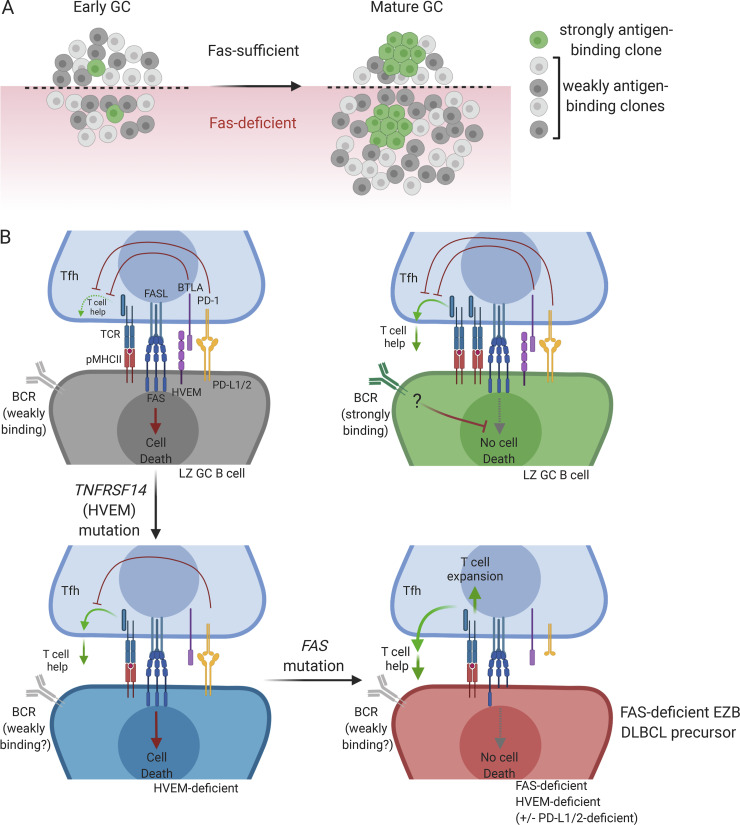
Fas is highly expressed on germinal center (GC) B cells, and mutations of FAS have been reported in diffuse large B cell lymphoma (DLBCL). Although GC-derived DLBCL has better overall outcomes than other DLBCL types, some cases are refractory, and the molecular basis for this is often unknown. We show that Fas is a strong cell-intrinsic regulator of GC B cells that promotes cell death in the light zone, likely via T follicular helper (Tfh) cell-derived Fas ligand. In the absence of Fas, GCs were more clonally diverse due to an accumulation of cells that did not demonstrably bind antigen. FAS alterations occurred most commonly in GC-derived DLBCL, were associated with inferior outcomes and an enrichment of Tfh cells, and co-occurred with deficiency in HVEM and PD-L1 that regulate the Tfh-B cell interaction. This work shows that Fas is critically required for GC homeostasis and suggests that loss of Tfh-mediated counterselection in the GC contributes to lethality in GC-derived lymphoma.
This is a work of the U.S. Government and is not subject to copyright protection in the United States. Foreign copyrights may apply.
Conflict of interest statement
Disclosures: N. Kotlov reported a patent to BostonGene issued. O. Plotnikova reported "BostonGene employee." G.A. Smith reported personal fees from AMGEN outside the submitted work. J.J. O'Shea reported "other" from Pfizer outside the submitted work. J.D. Phelan reported a patent to PCT/US2018/025377 pending. D.W. Scott reported personal fees from Abbvie, personal fees from AstraZeneca, personal fees from Celgene, personal fees from Janssen, grants from Janssen, and grants from NanoString outside the submitted work; in addition, D.W. Scott had a patent for molecularly subtype lymphoma pending with NanoString. No other disclosures were reported.
Figures
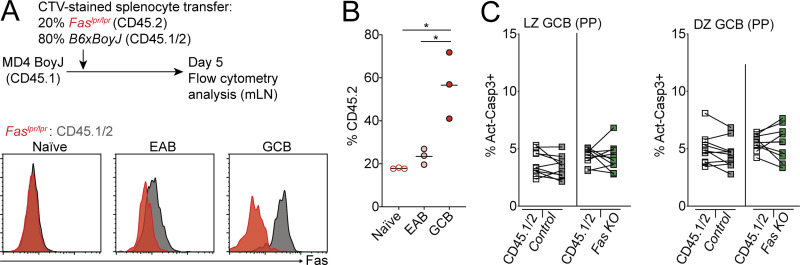
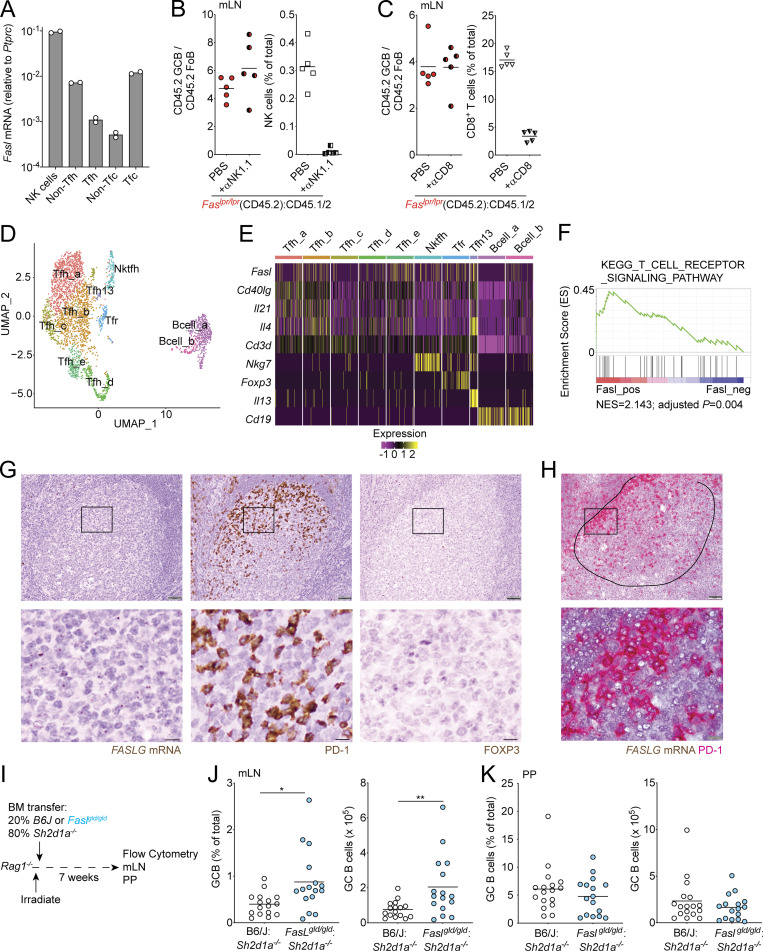
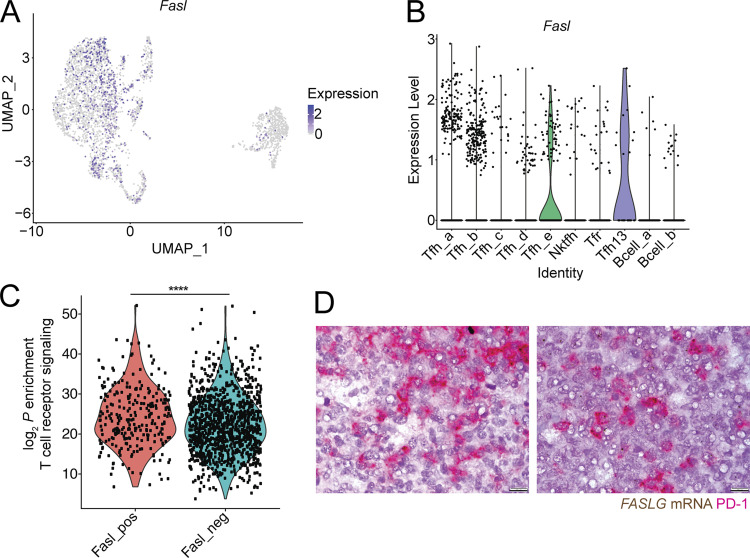
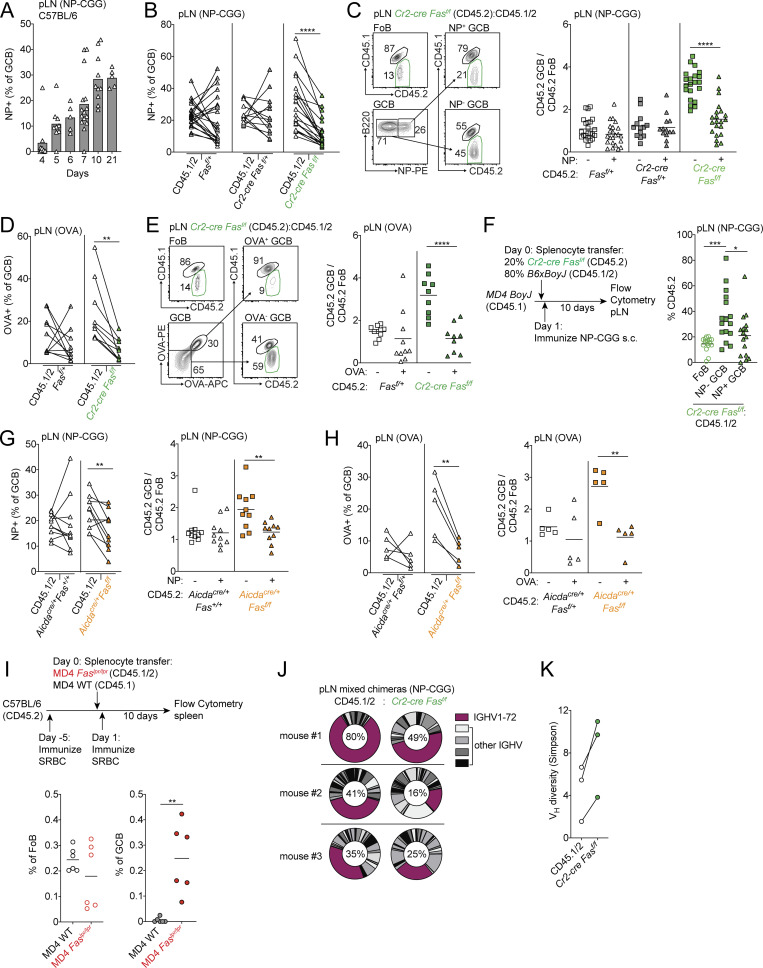
References
-
- Afshar-Sterle, S., Zotos D., Bernard N.J., Scherger A.K., Rödling L., Alsop A.E., Walker J., Masson F., Belz G.T., Corcoran L.M., et al. 2014. Fas ligand-mediated immune surveillance by T cells is essential for the control of spontaneous B cell lymphomas. Nat. Med. 20:283–290. 10.1038/nm.3442 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
